
Тип инфузории строение
Тип Инфузории строение фото простейшие животные ядро клетки рисунок вакуоль органоиды
Латинское название Ciliophora или Infusoria
Тип инфузории - высокоорганизованные одноклеточные с наиболее сложной системой органелл. Они характеризуются наличием двигательных органелл — ресничек, ядерным дуализмом и особой формой полового процесса — конъюгацией.
По современной классификации Тип/Отдел Ciliophora • 8,613 современных видов
Класс Oligohymenophorea - Инфузории 2,648 современных вида
Общая характеристика
Тип инфузории объединяет большое количество видов (свыше 6000) наиболее высокоорганизованных простейших.
Для них характерно присутствие ресничек, имеющихся обычно в большом числе. Реснички служат органеллами движения, они могут слипаться вместе, образуя более сложно устроенные органеллы. У некоторых сосущие инфузории реснички имеются только на ранних стадиях жизненного цикла. Для всех инфузорий характерен ядерный дуализм, т. е. двойственность. Это означает, что они имеют не менее двух ядер, различающихся как по размеру, так и по функции. Одно из ядер, значительно более крупное, называется макронуклеусом, а второе, маленькое — микронуклеусом. Некоторые виды инфузорий имеют по нескольку микро- и макронуклеусов. Микронуклеус служит половым, или генеративным, ядром, играющим основную роль в половом процессе. Макронуклеус — соматическое, или вегетативное, ядро, регулирующее все жизненные процессы, кроме полового процесса.
Бесполое размножение инфузорий происходит путем поперечного де-ления. Половой процесс у инфузорий протекает своеобразно, в виде конъюгации, которая не наблюдается у простейших других классов. Конъюгация заключается во временном сближении двух особей и взаимном обмене частями их микронуклеусов.
Инфузории — обитатели главным образом пресных водоемов, но встречаются также в солоноватой воде и в морях, некоторые виды приспособи-лись к существованию во влажной почве. Среди инфузорий много парази-тов (около 1000 видов) беспозвоночных и позвоночных животных.
Класс иразделяется на два класса:
- Ресничные инфузории (Ciliata);
- Сосущие инфузории (Suctoria).
Тип инфузории Класс ресничные
Латинское название Ciliatas
Ресничные инфузории составляют подавляющее большинство класса.. По образу жизни и способу питания они довольно разнообразны. Одни из них ведут плавающий образ жизни, другие обитают на дне или держатся на водных растениях. Большинство ресничных инфузорий питается бактериями и мелкими водорослями, однако есть и инфузории-хищники, поедающие других простейших, в том числе и других инфузорий. Среди ресничных инфузорий немало паразитов. Таковы инфузории, паразитирующие на коже рыб или в кишечнике различных животных и человека.
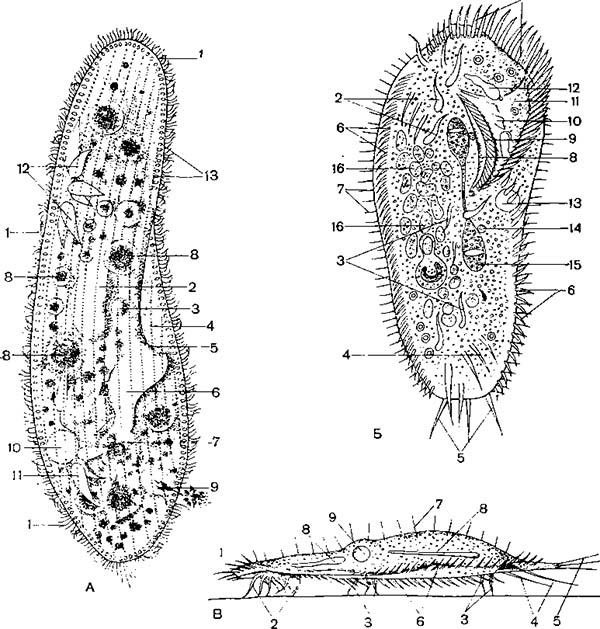
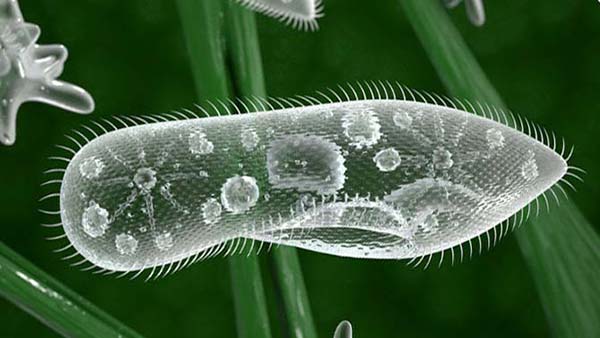
А — обыкновенная туфелька (Paramecium caudatum); 1— реснички; 2 — макронуклеу; 3— микронуклеус; 4— перистом; 5 — рот; 6 — глотка; 7 — образование пищеварительной в,акуолн; 8 — пищеварительные вакуоли; 9 — дефекация; 10 — резервуар сократительной ваку !>ли; 11, 12 — приводящие каналы сократительных вакуолей; 13 — трихоцисты; Б — брюхоре сннчная Stylonichia mytilus; 1 — адоральные мембранеллы; 2, 3, 4 и 5—группы лобных, брюшных, анальных и хвостовых цирр; 6 — ряд маргинальных цирр; 7 — спинные щетинкн; 8 — край перистома; 9 — иреоральные реснички; 10 — волнообразная перепонка; 11 — перистом; 12 — приводящий канал сократительной вакуоли; 13 — резервуар сократительной вакуоли; 14 — микронуклеус; 15 — макронуклеус; 16 — пищеварительная вакуоля; В — ползающая стилоннхия; 1 — адоральные мембранеллы; 2, 3, 4 и 5 — лобные, брюшные, анальные и XBOCI овые цпрры; 6 — маргинальные цирры; 7 — спинные щетинки; 8 — приводящие каналы; 9 — Сократительная вакуоля.
Инфузории имеют довольно разнообразную форму тела. Однако у многих видов в связи с приспособлением их к плавающему образу жизни форма тела удлиненная, обтекаемая. Примером может служить обыкновенная туфелька (Paramecium caudatum) (рис. 2, А). Размеры также различны, некоторые виды достигают довольно крупных размеров, до 2 мм длины (Spirostomum).
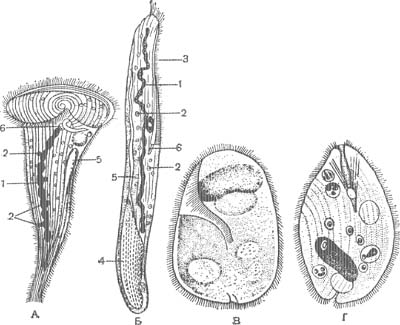
Тело ипокрыто тонкой, но прочной оболочкой — пелликулой, имеющей довольно сложную структуру. Пелликула гибка и эла-стична, поэтому она не служит препятствием для некоторого изменения ЛЬормы тела. Многие инфузории могут изгибать его, протискивать между различными предметами. У крупной инфузории «трубача» (Stentor) (рис. 43, А) тело вытянуто в виде граммофонной трубы, но оно может сильшЬ сжиматься и принимать шарообразную форму.
(/Р*снички — органеллы движения инфузорий. Они представляют собой очень тонкие и короткие многочисленные плазматические волоски. Ультг4атонкое строение ресничек и жгутиков, изученное с помощью* электронного микроскопа, показало их поразительное сходство.
У одних инфузорий реснички равномерно покрывают все тело. Например, у туфельки около 10 000—15 000 ресничек, расположенных правильными рядами. У других реснички сосредоточены в опреде-ленных местах тела.'Колебания ресничек представляют, по существу, гребные движения, состоящие из удара назад, при котором ресничка быстро движется в одной ш^кости, и возвращения в исходное положение, когда ресничка медленно двйк&тся вперед, плавно описывая полукруг. При комнатной температуре решшчки совершают около 30 взмахов в секунду. Движения ресничек происходят согласованно, в результате чего получаются правильные волнообразные колебания всех рядов ресничек. Туфелька движется со скорость до 2,5 мм/сек, т. е. за секунду проходит расстояние, в 10—15 раз превышающее длину ее тела.
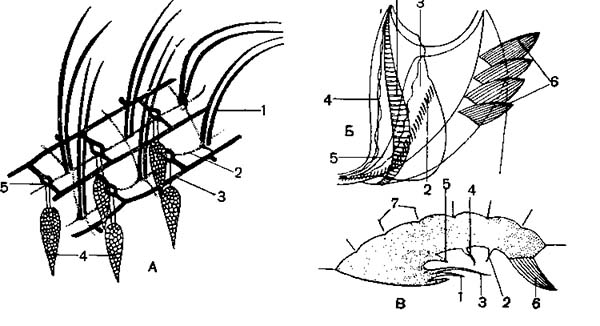
А — строение поверхности тела Paramecium nephridiatum; 1 — попарно сидящие реснички; 2 — невроплазматическая сеть; 3 — ребрышки пелликулы; 4 — трихоцисты; 5 — отверстие трихоцисты- Б ресничный аппарат перистома стилонихии (Stylonichia mytilus) с брюшной стороны; Вто же в поперечном разрезе; 1 — предротовые реснички; 2 — ротовые реснички; 3 — предротовая волнообразная перепонка; 4 — внутренняя волнообразная перепонка; 5 — ротовая волнообразная перепонка; 6 — мембранеллы; 7 — спинные щетинки.
Кроме простых ресничек, у них имеются более крупные образования, обычно окружающие ротовое углубление или расположенные на других частях тела. Это так называемые мембранеллы (рис. 2, Б). Каждая мембранелла представляет собой ряд ресничек, слипшихся вмсте в одну пластинку, часто имеющую треугольную форму (рис. 3, Б), если слипается более длинный ряд ресничек, образуется волнообразная /пере-понка, или мембрана. Такие перепонки имеются у многих в ротовом углублении или в глотке. Строение ресничного аппарата и рас-положение различных ресничных образований служат важными система-тическими признаками.
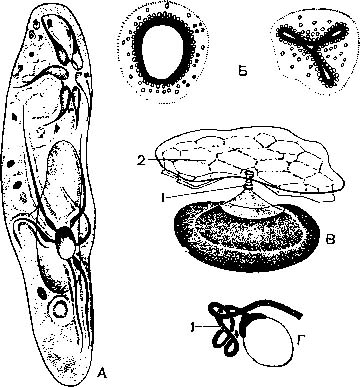
Цитоплазма ресничных отчетливо разделяется на наружный, более светлый и плотный слой — эктоплазму и более жидкий и зернистый внутренний слой — эндоплазму (рис. 2).
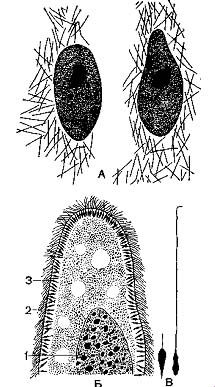
Эктоплазма имеет сложное строение, образуя большое количество органелл. Она выделяет на своей поверхности уже упоминавшуюся ранее эластичную пелликулу. У туфельки пелликула имеет сложную скульптуру: она образована правильными шестигранниками, в центре которых по-мещаются реснички. По-видимому, подобное строение увеличивает прочность наружной оболочки. Эктоплазме также принадлежат реснички и мембранеллы вместе с базальными тельцами. В эктоплазме многих ин-фузорий расположены в большом количестве так называемые трихоцисты (рис. 4). Это удлиненные палочковидные тельца, сильно преломляющие свет. При раздражении трихоцисты выбрасываются через особые канальцы наружу в виде тончайшей струйки жидкости, застывающей в воде тонкой упругой нитью. Трихоцисты — органеллы нападения и защиты. Хищные при помощи трихоцист парализуют добычу; «мирные» — защищаются от нападения хищников. По происхождению трихоцисты представляют собой видоизменение двигательных органелл и образуются из базальных телец.
В эктоплазме И. при соответствующей обработке можно обнаружить сеть тончайших волоконец, лежащих вблизи базальных телец и трихоцист (рис. 3, А). Полагают, что эти волоконца — нейрофаны — проводят раздражения и обусловливают согласованную работу ресничного аппарата. Однако во многих случаях подобные волоконца имеют опорное значение. Выше было указано., что многие из них могут изменять форму тела. Это обусловлено тем, что в эктоплазме расположены особые со-кратительные нити, или мионемы. Так, у трубача (Stentor) и некоторых других система сократительных мионем состоит из множества продольно расположенных волоконец, идущих вдоль тела и выстилающих околоротовое углубление (рис. 5 А). Наибольшей степени сложности достигает система мионем у Caloscolex из желудка жвачных, описанная проф. В. А. Догелем (рис. 5, Б). У сидячих инфузории сувоек имеется довольно сложна устроенный стебелек, внутри которого также проходят мионемы. При раздражении сувоек их стебелек свертывается в спираль (рис. 45).
Определенная форма тела, иногда довольно причудливая, обусловлена присутствием в эктоплазме плотных скелетных образований. Чаще всего это целая система опорных волоконец (рис. 5, В).
Таким образом, эктоплазма достигает высокой степени дифференцировки и содержит большое количество органелл: ресничный аппарат, трихоцисты, мионемы, нейрофаны и опорные скелетные волокна. В эндоплазме также расположены целые системы органелл: пищеварительных и выделительных (частично помещающихся, однако, и в эктоплазме).
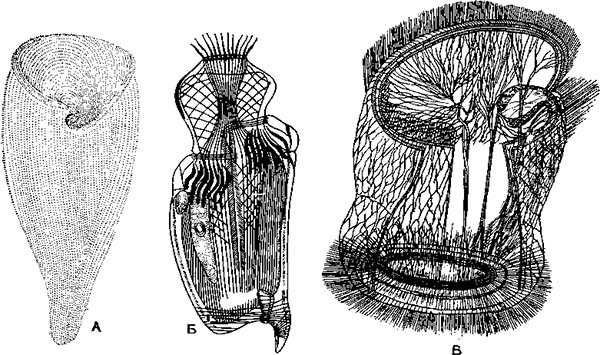
А — мионемы трубача (Stentor); Б — сложная система мионем у Caloscolex; В — система опорных волоконец у Trichodina pediculus, паразитирующей на гидрах.
Пищеварительные органеллы инфузорий начинаются ртом, или цитостомо м, представляющим собой отверстие в пелликуле. У многих рот помещается на дне особого углубления — околоротовой впадины, или перистома (рис. 2, А). У многих , питающихся мелкими организмами (бактериями), перистом окружен спирально расположенным венчиком мембранелл (разноресничные и кругоресничные). В перистоме может быть расположена волнообразная перепонка (рис. 2 и 3, Б).
Мерцательные движения ресничек и мембранелл вызывают токи воды, которыми пищевые частицы (бактерии и пр.) подносятся ко рту. У многих хищных перистома нет, и пищу они заглатывают сильно растягивающимся ртом (рис. 40, В).
Рот ведет в «глотку», или цитофаринкс, представляющую собой короткий канал, иногда также выстланный ресничками. У внутреннего края глотки образуется пузырек, состоящий из капельки жидкости, выделяемой эндоплазмой, в которую попадают накапливающиеся на дне глотки пищевые частицы. Так образуется пищеварительная вакуоля (рис. 2, А).
У туфельки при изобилии пищи примерно каждую минуту образуется новая пищеварительная вакуоля. Содержащие пищу вакуоли отрываются от глотки и перемещаются в эндоплазме инфузории, совершая определенный путь. Так, у туфельки каждая пищеварительная вакуоля сначала описывает малый круг в задней половине тела, а затем большой круг, доходя до переднего конца тела.
<
Процесс заглатывания, образование пищеварительных вакуолей и их передвижение в эндоплазме легко наблюдать при прибавлении в капельку воды с инфузориями растертой туши или кармина. Во время передвижения в вакуоле происходит переваривание пищи и всасывание переваренной пищи в эндоплазму. В пищеварительные вакуоли эндоплазма выделяет ферменты.
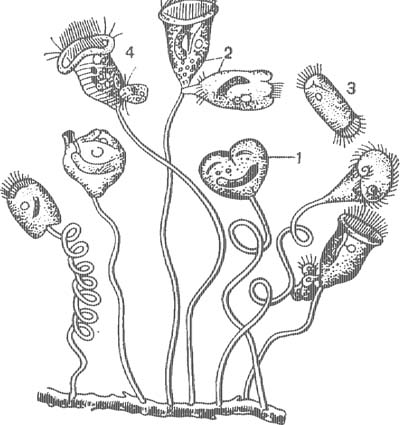
А — Bursaria truncatella; Б — Dileptus unser; В — Spathidium spatula; Г — Didinium, пожирающие туфельку.
Установлено, что на разных этапах пищеварения кислотность содержимого вакуоли различна. Вначале содержимое вакуоли имеет кислую реакцию, затем щелочную.
Вакуоли, содержащие непереваренные остатки пищи, подходят к поверхности эктоплазмы. У многих инфузорий на определенном месте тела, ближе к заднему концу, в пелликуле имеется особое отверстие — цитопрокт, через которое и совершается дефекация (рис. 2, А). Процесс дефекации происходит значительно реже процесса образования пищеварительных вакуолей (через 7—10 мин), так как перед дефекацией несколько вакуолей с непереваренными остатками пищи сливаются в одну. Весь процесс пищеварения у туфельки, от образования вакуолей до дефекации, длится в зависимости от температуры от 1 до 3 ч.
Как было сказано выше, среди инфузорий немало хищников, питающихся другими (рис. 6). Например, крупная хищная Bursaria заглатывает туфелек и других, загоняя их в глотку движением мембранелл. У других хищников заглатывание происходит иначе. Рот их сильно растяжим, и они заглатывают и втягивают довольно крупных инфузорий. Некоторые хищные могут поедать инфузорий, значительно превышающих их собственные размеры. Так, сравнительно небольшие Didinium (рис. 40, Г) нападают на туфелек, убивают их особым хоботком, затем постепенно втягивают и переваривают.
Выделительные органеллы представлены у них одной, двумя или несколькими сократительными вакуолями, расположенными в определенных частях тела (рис. 2). Сократительные вакуоли часто имеют довольно сложное строение (рис. 7). Помимо самой вакуоли, периодически сжимающейся (состояние систолы) и расширяющейся (диастолы), к ней ведут расположенные в эндоплазме приводящие каналы. Благодаря этому выделяющиеся вещества поступают в сократительную вакуолю из различных частей тела инфузории. От вакуоли к пелликуле ведет выводной проток, открывающийся особым отверстием наружу (рис. 7).
А — сократительные вакуоли и приводящие каналы Paramecium caudatum; Б — сократи¬тельные вакуоли Campanella umbel- laria в состоянии диастолы (слева) и систолы (справа); В — схема строения сократительной вакуоли Cycloposthium; вакуоля открывается наружу постоянным каналом, окруженным осо¬быми мионемами-замыкателями (2); 2 — пелликула; Г — сократительная вакуоля Paramecium trichium с извитым выводным каналом (2).
При наличии двух вакуолей (например, у туфельки) они сокращаются поочередно. При 16°С каждая вакуоля сокращается через 20—25 сек (у туфельки).
Инфузории, подобно другим простейшим, способны реагировать на разнообразные внешние раздражения. В отличие от многих жгутиковых инфузории не имеют светочувствительных органелл. Роль чувствительных органелл играют главным образом реснички и мембранел- лы. У одних реснички сохраняют при этом двигательную функцию; у других же, например у стилонихии, спинные реснички служат только осязательными органеллами.
Реакция на раздражение выражается в замедлении или ускорении, а также в изменении направления движения (туфельки), в свертывании перистома и сжатии тела (стенторы, сувойки), в сокращении стебелька
(сувойки) и т. п. Инфузории весьма чувствительны к малейшему прикосновению посторонних пре-метов. Они очень чувствительны и к изменению химического состава среды, причем различные вещества действуют на них по-разному, вызывая либо положительную, либо отрицательную реакцию. Способность различно реагировать на разные химические вещества имеет большое значение в жизни инфузорий при нахождении необходимой им пищи и наиболее благоприятных условий существования. Для дыхания оний необходимо достаточное количество растворенного в воде кислорода. Они,
как и другие простейшие, дышат всей поверхностью тела. Поэтому инфузории положительно реаги-руют на попадание в капельку воды пузырька воздуха, собираясь возле него. Инфузории реагируют положительно или отрицательно на изменение температуры среды, причем каждый вид характеризуется приспособленностью к определенной оптимальной для него температуре.
Ядерный аппарат ресничных инфузорий, как уже сказано, состоит из одного или нескольких макронуклеусов, имеющих различную форму (рис. 2 и 43), и одного или нескольких микронуклеусов. В деталях строение ядерного аппарата сильно варьирует. Так, обыкновенная туфелька (Paramecium caudatum) имеет один крупный макронуклеус и один микронуклеус, помещающийся в углублении макронуклеуса. У другого вида этого же рода — Р. aurelia — имеются два микронуклеуса. У сувоек макронуклеус имеет подковообразную форму, а у трубача, кроме очень удлиненного четковидного макронуклеуса, есть несколько микронуклеусов (рис. 43). Дифференцировка ядерного аппарата на вегетативное ядро — макронуклеус и на половое, или генеративное, ядро — микронуклеус характерна для всех ресничных инфузорий.
Микронуклеус отличается от макронуклеуса не только размером, но и числом хромосом. В то время как микронуклеус обладает диплоидным набором хромосом, макронуклеус полиплоиден, т. е. набор хромосом повторен у него много раз. Так, у туфельки Paramecium caudatum макронуклеус является 80-плоидным (по другим данным, 160-плоидным), а у близкого вида Р. aurelia — 1000-плоидным. У некоторых степень плоидности может доходить до 10—15 тыс.
Таким образом, ресничные инфузории по сравнению с другими простейшими имеют очень сложное строение. Оно усложняется в двух направлениях. Мы видели, что у инфузорий имеется большое число различных органелл, часто образующих целые системы, например систему пище-варительных, выделительных органелл и т. п. С другой стороны, для инфузорий характерно умножение, или полимеризация, многих органелл. Несомненно, реснички с базальными тельцами по происхождению соответствуют жгутиковому аппарату жгутиковых. Но по сравнению с полимеризацией локомоторных органелл у многожгутиковых, у инфузорий полимеризация идет значительно дальше. Развивается сложная система органелл, состоящая из огромного количества ресничек, частью превращающихся в мембранеллы, цирры и т. п. При этом сложность организации выражается в согласованном функционировании всего двигательного аппарата. Для инфузорий характерно также умножение числа ядер. Они имеют не менее двух ядер. Однако, в отличие от многожгутиковых, этот процесс усложняется еще дифференцировкой ядер.
Сосущие инфузории Suctoria
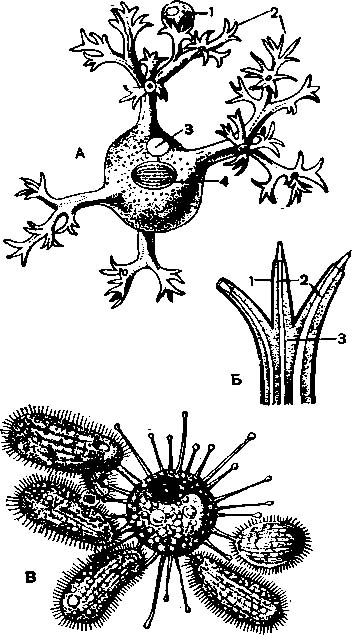
А — сосущая Dendrocometes para¬doxus; 1 — пойманная добыча; 2 — разветв¬ленные щупальца; 3 — сократительная вакуоля; 4— макронуклеус; Б — сосательное щупальце Dendrocometes; 1— пеллику¬ла; 2— канальцы; 3— цитоплазма; В— Sphaerophrya, сосущая нескольких ресничных.
Сосущие инфузории — небольшая специализированная группа хищников и паразитов, включающая несколько десятков видов. Все сук- тории — сидячие животные, в связи с чем во взрослом состоянии полностью лишены ресничек. Они прикрепляются различным образом к суб-страту, часто к покровам животных, например ракообразных, при помощи особой ножки, или подошвы.
У сосущих инфузорий отсутствуют обычные для ресничных органеллы: перистом, рот, глотка, пищеварительные вакуоли и т. п. Питание происходит при помощи особых сосательных щупалец, представляющих собой плазматические трубочки с каналом, ведущим внутрь эндоплазмы (рис. 46). Стоит какой-либо ресничному виду или жгутиконосцу случайно прикоснуться к сосательному щупальцу, как это простейшее прилипает к нему и оказывается в плену у сосущей инфузории. Последняя подтягивает к жертве другие щупальца. Оболочка простейшего растворяется в местах прикрепления щупалец, и цитоплазма жертвы переливается по трубочкам внутрь, в эндоплазму сосущей инфузории.
Сосательные щупальца могут быть расположены различно: пли радиально (Sphaerophrya, рис. В), или пучками на свободном конце сидящей инфузории, или на особых выростах — «руках» (Dendrocometes paradoxus; рис. А, Б). Сосущие живут и в пресных водах и в морях. Одни из них — хищники, поедающие других инфузорий; другие паразитируют внутри них. На-пример, Sphaerophrya sol паразитирует внутри туфельки.
Бесполое размножение сосущих инфузорий происходит путем наружного или внутреннего почкования. От материнской особи отделяются почки, снабженные пояском ресничек. Их называют бродяжками. Некоторое время они плавают, затем прикрепляются к субстрату, теряют реснички и превращаются в типичных сосущих инфузорий.
У сосущих имеется макронуклеус и микронуклеус. Половой процесс, так же как у ресничных, происходит путем конъюгации.
Наличие у бродяжек ресничек и двух ядер, а у взрослых сосущих полового процесса путем конъюгации сближает их с ресничными инфузориями и позволяет считать, что сосущие произошли от ресничных.
Поделиться с друзьями