
Класс ресничные инфузории
Латинское название Ciliatas
Класс Инфузории - Ciliophora или Infusoria
Общая характеристика
Класс ресничные инфузории - Обширный класс простейших. Имеют разнообразную форму, но чаще они овальные. Как и для других инфузорий, важным и общим признаком является присутствие в теле двух качественно различных ядер: крупного вегетативного ядра — макронуклеуса и гораздо более мелкого генеративного — микронуклеуса. В течение всей жизни тело покрыто ресничками (за исключением стадий инцистирования). В эктоплазме залегают особые защитные приспособления — трихоцисты — короткие палочки, которые при раздражении животного выстреливают наружу, превращаясь в длинную упругую нить. Нити вонзаются в тело врага или добычи и парализуют его. Как и у других инфузорий, имеется сложная система органелл пищеварения. Свободноживущие инфузории встречаются как в пресных водах, так и в морях, могут жить в почве. Ярким представителем является инфузория туфелька. Питаются бактериями, одноклеточными водорослями. Инфузории-хищники питаются другими инфузориями, ресничными червями. Среди инфузорий есть симбионты жвачных, которые обитают в переднем отделе их желудка (рубце и сетке). Многочисленной группой являются паразитические инфузории, паразитирующие на коже (Ichthyophthirius) и жабрах (Trichodina) рыб, толстом кишечнике чело-Iвека (Balantidium coll).
Размножение
Бесполое размножение ресничных инфузорий происходит путем поперечного деления их тела надвое, которое начинается с деления обоих ядер. Первым делится микронуклеус митотическим путем. После этого макронуклеус вытягивается в длину, затем образуется перетяжка, и он делится. Раньше считали, что макронуклеус делится прямым делением, или амитотически. Исследования последних лет открыли высокую степень полиплоидностп макронуклеусов и сложные процессы их перестройки при каждом делении. Этот тип деления высокополиплоидных ядер, внешне напоминающий амитоз, назван сегрегацией геномов (их разъединением).
В это же время на теле инфузории появляется постепенно углубляющаяся поперечная перетяжка и две образовавшиеся дочерние особи отделяются одна от другой. При этом происходит глубокая внутренняя перестройка дочерних инфузорий. В них не только формируются недостающие органеллы, но и обновляются многие органеллы, присутствующие у обеих особей (старые резорбируются, а новые возникают). Вазальные тельца ресничек делятся, после чего формируются новые реснички. Весь процесс деления у туфельки продолжается от 30 мин до 2—3 ч в зависимости от температуры.
Бесполое размножение у туфелек происходит регулярно 1—2 раза в сутки, при повышении температуры частота делений увеличивается. Некоторые крупные инфузории, например трубач, делятся значительно реже, один раз в несколько дней. После ряда бесполых размножений наступает половой процесс.
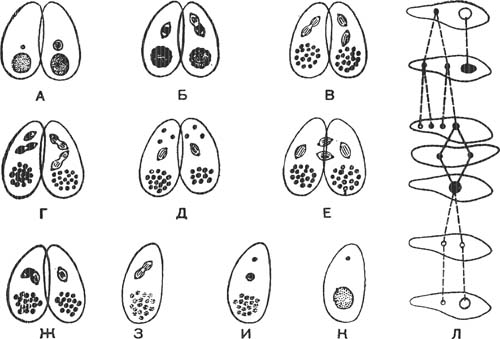
А — начало конъюгации; Б и В — первое деление микронуклеуса; Г — второе деление микронуклеуса; Д — начало третьего деления в одном из четырех микронуклеусов каждого конъю- ганта; Е — обмен мигрирующими пронуклеусами; Ж — слияние пронуклеусов; 3, И, К — деление синкариона и образование нового ядерного аппарата; Л — схема аутогамии.
Половое размножение у инфузорий связано с временным попарным соединением особей и носит название конъюгации. В типичном случае (например, у Paramecium caudatum) половой процесс происходит следующим образом (рис. 2). Две инфузории прикладываются друг к другу брюшными сторонами, на которых расположены перистомы с ротовыми отверстиями. В определенных местах пелликула растворяется, и конъюганты оказываются соединенными небольшим плазматическим мостиком.
В таком состоянии конъюгирующие особи плавают, и в это время происходит ряд изменений в каждом из конъюгантов. Макронуклеусы в половом процессе никакого участия не принимают. Они начинают постепенно дегенерировать и в конце концов распадаются на части, которые рассасываются в плазме. С микронуклеусами каждого из конъюгантов происходят сложные изменения: они дважды делятся, в результате чего получаются четыре ядра, из которых три рассасываются, а одно остается. Эти два деления микронуклеуса соответствуют двум делениям созревания половых клеток многоклеточных животных. Одно из делений ядра — ре-дукционное, в результате которого парные гомологичные хромосомы, не делясь, расходятся к разным полюсам ядра. При этом происходит р е- дукция, или сокращение числа хромосом в разделившихся микронуклеусах. Ядро из диплоидного становится гаплоидным. Оставшееся ядро еще раз делится митотически, в результате чего образуются два гаплоидных ядра, или пронуклеуса. Они отличаются друг от друга тем, что одно из них неподвижное — его называют стационарным ядром, а другое подвижное — мигрирующее ядро. Мигрирующие пронуклеусы обоих конъюгантов подходят к соединяющему их плазматическому мостику, и конъюганты обмениваются ядрами. Затем в каждом конъюган- те мигрирующее ядро соединяется со стационарным, образуя одно ядро — синкарион.
Слияние двух гаплоидных ядер, в результате которого восстанавливается диплоидный набор хромосом у синкариона, является процессом перекрестного оплодотворения. Его можно сравнивать с процессом оплодотворения у многоклеточных. Сущность оплодотворения у многоклеточных состоит в слиянии их половых клеток, или гамет. У инфузорий гаметы не формируются, но зато образуются половые ядра. Стационарное ядро иначе называют женским. У некоторых инфузорий оно отличается более округлой формой от мигрирующего, или мужского, ядра, которое может быть похожим на сперматозоид (рис. 3, В).
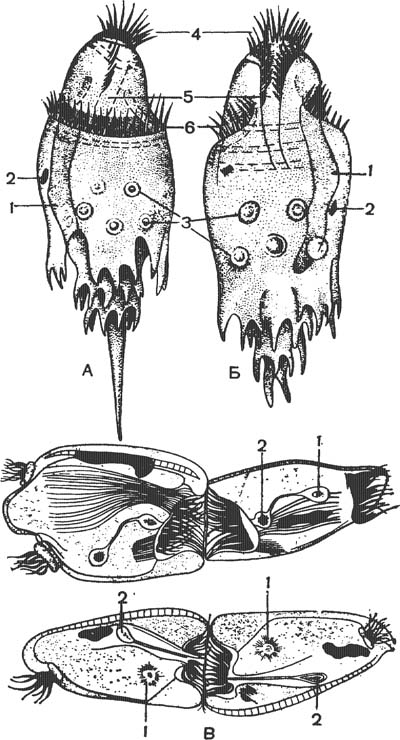
А — Ophryoscolex caudatus; Б — О. purkinjer; 1 — макронуклеус; 2 — микронуклеус; 3 — сократительные вакуоли; 4 — околоротовые мембранеллы; 5 — глотка; 6 — пояс мембранелл; В — две стадии конъюгации Cycloposthium bi-palmatum: вверху — деление микронуклеуса на мигрирующее (2) и стационарное (i) ядра, внизу — обмен мигрирующими (мужскими) ядрами.
При конъюгации происходит слияние только ядер — мужского и женского, а не клеток, другими словами, имеет место только кариогамия, или слияние ядер, а не гаметогамия (слияние гамет). Однако есть основание полагать, что вместе с мигрирующим ядром проникает и некоторая часть цитоплазмы. В итоге каждая из конъюгирующих инфузорий имеет заново сформированное ядро — синкарион. У инфузорий, имеющих два и больше микронуклеусов, в образовании синкариона, по-видимому, принимает участие только один микронуклеус каждого конъюганта. На этом конъюгация заканчивается, и конъюганты расходятся. Длительность процесса конъюгации от начала спаривания до расхождения конъ- югантов у туфельки достигает 12 ч, некоторые инфузории остаются спаренными несколько суток.
Восстановление ядерного аппарата (формирование новых макро- и микронуклеусов) происходит в каждом эксконъюганте после их расхождения. После этого в каждой инфузории (в простейшем случае) синкарион делится надвое и одно ядро развивается в новый макронуклеус, а другое — в микронуклеус. Через некоторое время инфузории переходят к обычному размножению делением. Однако у туфельки до начала размножения синкарион делится последовательно не на 2, а на 8 ядер, из которых 4 более крупных — макронуклеусы, а 4 более мелких — микронуклеусы. Из четырех микронуклеусов три рассасываются, и в инфузории остается, таким образом, 4 макронуклеуса и 1 микронуклеус. В таком случае следующие за этим первые два деления инфузорий носят особый характер: при первом и втором делениях делится только микронуклеус, тогда как 4 макронуклеуса без деления распределяются по дочерним клеткам.
Таким образом, при половом процессе (конъюгации) происходит полная перестройка тела инфузории. Старый макронуклеус, играющий большую роль во всех процессах жизнедеятельности инфузорий, распадается; синкарион возникает после слияния мужского и женского ядер двух раз-личных особей, и новые макронуклеусы и микронуклеусы образуются из синкариона. Несомненно, происходят глубокие изменения и в плазме инфузории. Во время конъюгации цитоплазма конъюгантов сообщается через цитоплазматический мостик, вместе с обменом ядра происходит и обмен частью цитоплазмы.
Многочисленные исследования, проведенные над простейшими, показали, что если после каждого деления отсаживать инфузории Paramecium aurelia поодиночке, исключая, таким образом, возможность конъюгации, то можно получить очень большое количество бесполых поколений инфузорий (15 000 поколений). Оказалось, что при невозможности конъюгации в культуре инфузорий периодически обязательно происходит (через 25—30 бесполых поколений) перестройка ядерного аппарата, в основном аналогичная той, которая наблюдается при конъюгации.
Это явление получило название аутогамии, и заключается оно в следующем (рис. 2, Л). Макронуклеус распадается на части и рассасывается, микронуклеус делится на четыре ядра. У некоторых инфузорий три из этих ядер погибают, а одно делится еще раз на два пронуклеуса, которые затем сливаются вместе. Образовавшийся таким образом синкарион далее делится, образуя новый макронуклеус и новый микронуклеус. Процесс аутогамии считают соответствующим процессу самооплодотворения у многоклеточных.
У некоторых видов инфузорий реорганизация ядерного аппарата без конъюгации может проходить иначе. Так, у многих видов наблюдается разрушение и рассасывание макронуклеуса и одновременное деление микронуклеуса 2, реже 1 раз. Часть образующихся ядер резорбируется (рассасывается), а из 1—2 оставшихся восстанавливаются макро- и микронуклеусы без их предварительного слияния, т. е. без кариогамии. Эта форма реорганизации ядерного аппарата инфузорий названа эндо- мик исом.
Многочисленные исследования показали, что период бесполого размножения инфузорий можно значительно увеличить, если культивировать их в оптимальных условиях внешней среды, регулируя интенсивность питания, температуру и химический состав культуральной жидкости и удаляя из нее продукты метаболизма (обмена веществ). Последние могут накапливаться в среде и отравлять культуру. Эти опыты показали, что наступление конъюгации или иных форм полового процесса (аутогамии и эн- домиксиса) зависит не только от внутренних генетических особенностей инфузорий, но и от условий внешней среды.
По-видимому, жизнеспособность разных видов инфузорий в разной степени зависит от периодичности конъюгации, эндомиксиса и аутогамии. У одних инфузорий (Р. aurelia и др.) существует четко выраженный жизненный цикл с обязательным чередованием бесполого размножения и полового процесса. Нарушение этого цикла путем препятствия половому процессу приводит инфузорий к депрессии и гибели. Другие инфузории могут длительное время размножаться только бесполым путем. Так, туфелька, по данным последних лет, оказалась неспособной к аутогамии. По- видимому, она может размножаться бесполым путем неограниченно долго при оптимальных условиях ведения культуры.
В природных условиях практически не бывает оптимального сочетания всех факторов среды. Поэтому бесполое размножение всех инфузорий, как правило, чередуется с присущим им половым процессом — конъюгацией. Половой процесс (на примере конъюгации инфузорий) дает преиму-щества обоим партнерам (эксконъюгантам) с большим успехом противостоять изменяющимся условиям окружающей среды. При слиянии генетически разнородных ядер — пронуклеусов — обоих конъюгантов повышается изменчивость их различных физиологических и морфологических признаков. Обновленный макронуклеус при этом по-новому программирует жизненные функции инфузории. Не нужно думать, однако, что у всех инфузорий после конъюгации наступает своего рода «омоложение», выражающееся в повышении их жизнеспособности. Действительно, потомки одних эксконъюгантов обнаруживают повышенный обмен веществ и чаще делятся, тогда как потомки других эксконъюгантов (они могут быть из одной и той же пары) могут снижать темпы деления и быстро погибать. Это можно объяснить неблагоприятным сочетанием генетического материала при перестройке ядерного аппарата (возникновением летальных комбинаций генов). В процессе естественного отбора эти инфузории погибают, зато выживают и увеличиваются в числе более жизнеспособные.
Ресничные инфузории Классификация
Подкласс ресничных инфузорий (Ciliata) разделяется на несколько отрядов. Классификация основана главным образом на строении ресничного аппарата.
Важнейшие отряды следующие:
- Равноресничные (Ноlotricha);
- Спиральноресничные (Spirotricha);
- Кругоресничные (Реritricha).
ОТРЯД 1. РАВНОРЕСНИЧНЫЕ (HOLOTRICHA)
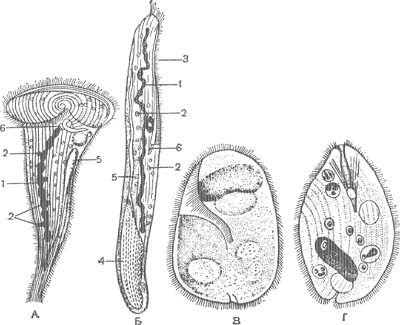
Этот самый большой и распространенный отряд инфузорий характеризуется наиболее примитивно устроенным ресничным аппаратом. У большинства равноресничных инфузорий все тело покрыто большим количеством ресничек, не образующих мембранелл. В области перистома реснички могут быть несколько длиннее. В глотке они иногда, слипаясь, образуют волнообразную перепонку. Типичными представителями равноресничных являются самые обычные виды рода Paramecium и др. У некоторых видов (род Didinium) реснички расположены не равномерно по всему телу, а поясками (рис.4, Г).
Большинство равноресничных питается бактериями, но есть среди них хищники и паразиты. Из хищников интересны уже упомянутые виды рода Didinium. Из паразитических равноресничных наибольшее значение имеют паразиты рыб., например Ichthyophthirius multifiliis, паразитирующий на коже и жабрах рыб. Взрослый паразит округлой формы, достигает больших размеров, до 1 мм в диаметре. Рыба, пораженная ихтиофтириусом, кажется посыпанной манной крупой. Крупные инфузории покидают тело рыбы, некоторое время свободно плавают, затем оседают на дно и инци- стируются. Под оболочкой цисты паразит многократно делится, в результате чего образуется около 2000 маленьких инфузорий. Они покидают цисту, попадают на кожу или жабры рыбы и активно проникают в ее живую ткань. Продолжительность паразитирования ихтиофтириуса в рыбе зависит от температуры. В жаркую погоду паразит дорастает до взрослой формы за 1— 2 недели, а в холодную развитие затягивается до 4—6 недель. Ихтиофтириус особенно опасен для мальков рыб, он может вызывать массовую гибель рыбьей молоди в рыбоводческих хозяйствах.
<
Спиральноресничные (Spirotricha);
А — Bursaria truncatella; Б — Dileptus anser; В — Spathidium spatula
Для представителей этого отряда наиболее характерно присутствие околоротовых мембранелл, расположенных по спирали, закрученной вправо.
Отряд спиральноресничных включает подотряды
- разноресничных (Heterotricha),
- брюхоресничных (Hypotricha)
- малоресничных (Oligotricha).
Подотряд 1. Разноресничные (Heterotricha).
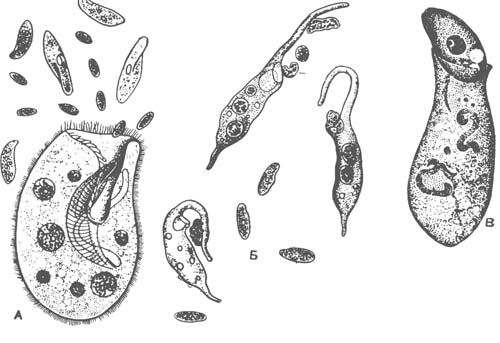
У разноресничных, кроме спирали крупных околоротовых мембранелл, имеются мелкие реснички, которые равномерно покрывают все тело. Типичными представителями разноресничных могут служить обычные в наших пресных водах инфузории — трубачи (Stentor). Они имеют своеобразный четковидный макронуклеус и несколько микронуклеусов. Трубачи могут временно прикрепляться суженным задним концом к водным растениям или грунту. Обладая хорошо развитой системой мионем, они способны в ответ на раздражение мгновенно изменять форму тела, превращаясь в почти правильный шар. Трубачи, так же как и некоторые другие роды разноресничных [(Spirostomum, Bursaria), являются одними из наиболее крупных инфузорий (рис. 5 А; 3), некоторые виды достигают 1—2 мм длины.
У разноресничных резко выражена способность к регенерации. Таких инфузорий, как Stentor или Spirostomum, можно разрезать на несколько кусков, причем каждый кусок регенерирует в целую инфузорию. Необходимо только, чтобы в отрезанной части инфузории была часть макронук-леуса и один из микронуклеусов.
6 — глотка.
Среди разноресничных имеются паразиты. Из них наибольшее значение имеет балантидий (Balantidium coli), опасный паразит кишечника человека (рис. 1, Г). К счастью, балантидий довольно редко встречается у людей, попадая к ним от своего основного хозяина — свиньи. Цисты балантидия заносятся в организм человека вместе с загрязненной пищей. В толстой кишке из них выходят инфузории, которые могут некоторое время обитать в просвете кишки, не причиняя вреда. Однако скоро балантидии внедряются в ткани, разрушая стенку толстой кишки и вызывая образование глубоких язв. Происходит интенсивное размножение паразитов, причем часть из них выходит в просвет кишечника, спускается в его нижний отдел, где и образует цисты. Цисты выводятся наружу вместе с фекальными массами и могут послужить источником дальнейшего распространения паразита.
Балантидиоз — тяжелое заболевание, проявляющееся как упорно протекающий колит с кишечными кровотечениями и приводящее к резкому малокровию. Лечение балантидиоза затруднительно из-за внутритканевой локализации паразита.
Подотряд 2. Брюхореспичные (Hypotricha).
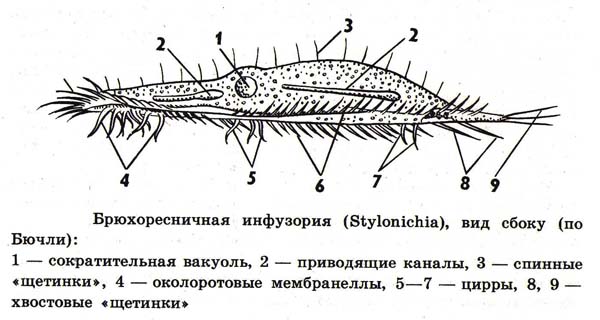
Брюхоресничные отличаются тем, что, кроме огибающих перистом мембранелл, они имеют предротовую волнообразную перепонку (рис. 1). Разбросанные по спинной стороне одиночные тонкие реснички малочисленны и почти неподвижны, они выполняют осязательную функцию. Тело брюхоресничных сплющено в спиннобрюшном направлении. На брюшной стороне имеются особые плотные щетинки — ц г склеившихся ресничек (рис. 4, В). по субстрату. Типичными представителями служат обычные в пресных водах стилонихия (Stylonichia mytilus), окситриха (Oxytricha), эуплотес (Euplotes) и др.
Подотряд 3. Малоресничные (Oligotricha).
У всех малоресничных наблюдается сильное сокращение ресничпого покрова. От него сохраняются только околоротовые мембранеллы, а у некоторых видов на теле имеются также отдельные реснички и цир-ры. Наиболее распространены своеобразные морские планктонные инфузории (Tintinnoidea), образующие легкие хитиноидные раковины.
Кроме того, к малоресничным относится своеобразная группа паразитических энтодиниоморфных, часто выделяемая в самостоятельный подотряд.
Энтодиниоморфные характеризуются сильной редукцией ресничного аппарата. У них имеется только околоротовая спиральная мембранелла и пояски или пучки мембранелл на теле (рис. 3). Типичными представителями энтодиниоморфных могут служить различные вида инфузорий из семейства Ophryoscolecidae, имеющие довольно причудливую форму тела с отростками, образуемыми пелликулой (рис.3, А, Б). Форма инфузорий обусловлена довольно сложной системой опорных элементов.
У представителей другого семейства—Cyclopostidae (рис. 3,В) — В. А. Догель обнаружил, что образующиеся при конъюгации мигрирующее (мужское) ядро отличается от стационарного тем, что окружено слоем плазмы, образующей подобие хвостика сперматозоида» При конъюгации инфузории соединяются между собой перистомами, через которые мигрирующие ядра проникают в цитоплазму партнера. Часть пути ядра проходят вне тела инфузории.
Энтодиниоморфные особенно часто встречаются в рубце и сетке желудка жвачных. «Нельзя найти ни одного экземпляра домашнего скота, кишечник которого не содержал бы обильной фауны этих инфузорий»,— писал В. А. Догель, специально занимавшийся исследованием этой группы инфузорий. По его данным, в 1 см3 содержимого рубца находится более 1000 000 инфузорий. Общая масса инфузорий в рубце одного жвачного животного составляет около 3 кг. Однако эти инфузории не приносят вреда своему хозяину, и их можно считать скорее симбионтами, чем паразитами. Употребляя в пищу клетчатку, имеющуюся в изобилии в пище жвачных животных, инфузории переваривают ее. Полагают, что эти инфузории способствуют размельчению, перевариванию и, в конечном счете, усвоению пищи их хозяевами — жвачными. Кроме того, большое количество инфузорий постоянно переваривается жвачными животными вместе с растительной массой, в результате чего животный белок тела инфузорий дополняет рацион их хозяев. Однако эти чрезвычайно важные для животноводства вопросы до конца не выяснены. Есть указания на то, что у ослабленных и больных животных энтодиниоморфные инфузории могут переходить к настоящему паразитированию.
Круглореснитчатые (PERITRICHA)
Кругоресничные инфузории в подавляющем большинстве ведут сидячий образ жизни. Ресничный аппарат представлен спиральным венчиком мембранелл, окружающих перистом. Спираль околоротовых мембранелл у кругоресничных закручена влево, в отличие от спиральноресничных, у которых она завертывается вправо. На остальной части тела реснички обычно отсутствуют. Только у плавающих стадий на заднем конце тела имеется венчик ресничек.
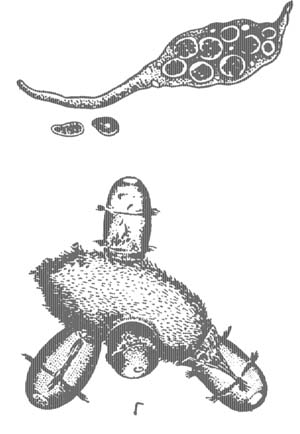
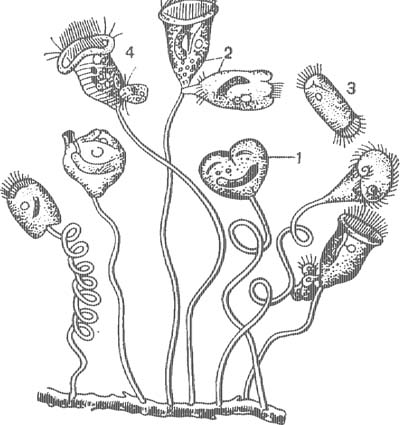
Из кругоресничных инфузорий в наших водоемах обычны сувойки (Vorticella), прикрепляющиеся к растениям, раковинам моллюсков длинными сократимыми стебельками, при сокращении свертывающимися в спираль (рис. 7). Тело сувоек также сократимо. При малейшем раздражении перистомальное поле с мембранеллами втягивается внутрь и инфузории принимают шаровидную форму. В отличие от одиночных сувоек (Vorticella) виды другого рода — Carchesium — колониальные. Колониальны также встречающиеся в пресных водах виды рода Zoothamnium, образующие зонтикообразные колонии на листьях различных водных растений.
При бесполом размножении у сувоек наблюдается деление надвое, причем одна особь остается на стебельке, а другая, отделяющаяся особь образует на заднем конце тела венчик нежных ресничек (рис. 7, 3). Такие «бродяжки» отрываются от материнской особи, плавают, а затем при-крепляются к субстрату, образуя стебелек и теряя реснички. Перед конъюгацией, однако, отделяются особи значительно меньших размеров. Их называют микроконъюгантами. Микроконъюгаиты прикрепляются к обычным сидячим особям — макроконъюгантам (рис. 7, 4). В процессе конъюгации происходит перестройка ядерного аппарата обоих конъюгантов, но синкарион образуется только в макроконъюганте. Следовательно, вместо взаимного оплодотворения происходит лишь одностороннее оплодотворение макроконъюганта, а микроконъюгант погибает. Таким образом, в данном случае можно считать микроконъюганта мужской особью, а макроконъюганта — женской.
Поделиться с друзьями