
Радиолярии значение скелет фото
Радиолярии фото биология скелет строение значения простейшие питание представители тип
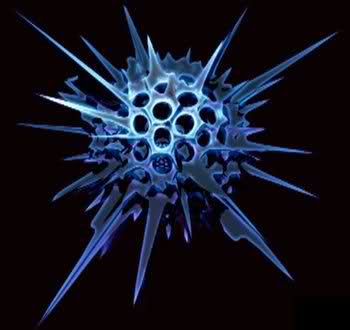
Внешний вид
Внешняя форма разнообразна. В большинстве случаев тело имеет форму шара и расходящиеся элементы скелета и псевдоподии. Размеры живой клетки радиолярий изменяются от нескольких десятков микрометров до 1000-3000 мкм. Редко радиолярии создают колонии, которые могут достигать нескольких метров. Окраска тела зависит от наличия зёрен пигмента и бывает красной, синей, зелёной и др.
Строение клетки
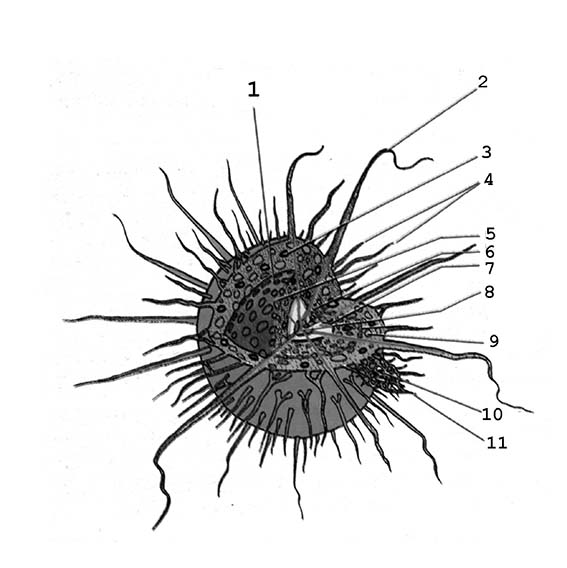
1 – слои эктоплазмы; 2 – аксоподии; 3 – зооаксантеллы; 4 – филоподии; 5 – ядро; 6 – эндоплазма; 7 – мембрана центральной капсулы; 8 - аксопласт; 9 – вакуоли; 10 – добыча; 11 - филлоподии
Минеральный скелет
Скелет у радиолярий исключительно внутренний, поэтому не подвергается растворению в течение жизни клетки.
Главное значение скелета для радиолярий в том, что они получают пассивную способность к парению в воде. Скелет радиолярий отличается большой прочностью и хорошо сохраняется в ископаемом состоянии, кроме целестиновых скелетов - они растворяются в морской воде после гибели организма.
- Простейший. Состоит из отдельных простых или двойных игл.
- Астроидный. Представлен двадцатью иглами радиально расходящимися от капсулы, но соединёнными в центре.
- Сфероидный. Противоположные иглы соединяются в центре и скелет оказывается состоящим из 10 игл, формируется более сложный шаровидный скелет.
Встречаются также скелеты в форме башенки, шлема, суженный на одном конце и расширенный на другом. Устьевый край скелета может закрываться решётчатой диафрагмой. При увеличении размеров
в области устья появляются пережимы, приводящие к образованию многокамерного скелета. Вершина таких скелетов называется цефалис. Головной отдел идущий за вершиной называется торакс (вторая камера). Грудной отдел (третья камера) называется абдомен. Последующие капсулы называются – постабдомен.
Формирование скелета
Завершение формирования скелета осуществляется двумя способами:
- способом мостика – формирование из прямых балок (перекладин), которые, соединяясь мостиками и разрастаясь, формирует скелет с полигональными порами;
- способом ободка - На органической матрице, которая быстро окремнеевает, а оставшиеся поры сужаются за счет утолщения стенки скелета.

Радиолярии Морфология скелета
Внутренний каркас
Значение внутренних каркасов еще остается не выясненным, поскольку подобная структура не сохранилась в скелете у современных полицистин, только у насселярий.
Типы внутреннего каркаса (по усложнению)
- Полая сфера – внутренний каркас представляет собой полую неперфорированную сферу с отходящими от нее полыми лучами. Не получил дальнейшего развития за счет массивности.
- Полусфера - тип внутреннего каркаса встречается крайне редко и только у отдельных ордовикских представителей.
- Многогранник - возможно, это первый шаг радиолярий на пути облегчения внутреннего каркаса скелета. Но и этот тип внутреннего каркаса не получил развития у других представителей радиолярий.
- Спикула – внутренняя первичная структура, морфологический центр скелета, представляет собой n-лучевую фигуру (где значение n равно от 4-6 до 24), образованную одной первичной четырехлучевой спикулой или объединением двух первичных четырехлучевых спикул. Первичная спикула чаще всего располагается внутри сферы в ее центре, но иногда спикула расположена эксцентрично. На дистальном окончании лучей спикулы формируются основные иглы скелета.
- Четырехлучевая спикула является универсальным первичным скелетным элементом радиолярий, устойчивым в пространстве и времени.
- Спикула со срединной балкой. Строение внутреннего каркаса в виде шестилучевой спикулы со срединной балкой, видимо, отвечало более нормальному функционированию клетки. Шести- и многолучевая спикула со срединной балкой образована из двух первичных четырехлучевых спикул. Этот тип спикулы характерен для сферических губчатых радиолярий, пористых представителей и радиолярий с пиломом.
- Микросфера - первичная микросфера развивается от углов спикулы. Микросфера связана с внешней оболочкой скелета иглами, являющимися продолжением спикулы, при этом одна игла - антапикальная - возникает от антапикальной части пер¬вичной микросферы.
Первичная внутренняя сфера
Отходящие от спикулы лучи в результате разрастания, ветвления и слияния дистальных концов образуют первичную внутреннюю сферу (Афанасьева, Амон, 2006). Диаметр первичной сферы современных радиолярий колеблется от 15 до 50 мкм и больше (Петрушевская, 1986). На основании соотношения диаметра внешней и первичной внутренней оболочки скелета можно выделить четыре типа первичной сферы: очень большая, большая, маленькая и очень маленькая. Стенка первичной сферы очень тонкая 1-3 мкм.
Оболочки скелета
- решетчатые, сетчатые и губчатые оболочки, скелетная ткань которых образована способом мостика;
- пористые и пластинчатые оболочки, в образовании скелетной ткани которых доминирует способ ободка.
2. Сетчатые оболочки. Стержни, формирующие сетчатые сферы, более тонкие, не шире, чем ячеи между ними, и связаны в целостную сеть. Сеть может быть одно- или многослойной. Углы ячей всегда сглажены, закруглены для увеличения прочности скелета.
3. Губчатые оболочки. Губчатые формы отличаются хаотичным переплетением наиболее тонких скелетных нитей, образующих спутанно-волокнистую структуру.
4. Пластинчатые оболочки. Неперфорированные пластинчатые оболочки известны достаточно широко, но иногда и в них есть, хотя бы редкие, полигональные поры.
5. Пористые оболочки. Наиболее широко распространенной, особенно у сферических полицистин, является пористая оболочка. Скелетная ткань пористой оболочки образована по способу ободка: края пор нарастают внутрь.
Скелет может быть пронизан порами одного или разного размера. Диаметр пор меняется от 1 до 35 мкм, но иногда встречаются очень тонкие пластины с порами 0,2-1 мкм. Расположение пор бывает беспорядочным, особенно если поры разной величины и формы, или упорядоченным, при этом оно часто напоминает шахматную доску. Иногда продольные или поперечные ряды пор вырисовываются более отчетливо, потому что стенки между рядами шире и/или толще, чем между порами в одном ряду. Кроме того, крупные поры могут быть затянуты тонкими ажурными сеточками или пористыми пластинками (Назаров, Петрушевская, 1996). Внешняя оболочка скелета может быть осложнена системой относительно высоких гребней, образующих ячеистую структуру поверхности скелета.
Устье и пилом
Иглы у радиолярий
Стержневидные иглы, как правило, очень тонкие и длинные с округлым поперечным сечением. Скелетные стержни могут быть непосредственно включены в стенку скелета, или могут быть укреплены у основания дополнительными перекладинами, создающими как бы очень широкие растяжки или подпорки.
Конусовидные иглы более прочные по сравнению со стержневидными. Они могут быть прямыми или изогнутыми и, так же как стержневидные, характеризуются округлым поперечным сечением игл. Однако в основании иглы иногда приобретают трехгранное строение за счет укрепления основания «ложными» растяжками.
Гранёные иглы обеспечивают ту же прочность, что и конусовидные, но при меньшем расходе материала. Наиболее выгодны иглы, состоящие из трех спаянных пластин вдоль одной грани.
Существуют с y-образным сечением игл. При этом иглы имеют лопастевидное сечение граней и могут быть тонкими, изящными или грубыми, очень массивными.
Наиболее оптимальной формой являются иглы с Y-образным сечением иглы и субпрямоугольным сечением граней. Форма граней у таких игл может быть субтреугольной, листообразной или узкой саблевидной.
Второстепенные иглы. Кроме основных игл, которые связаны с внутренним каркасом, у сферических радиолярий существуют второстепенные иглы, образующиеся на наружной поверхности скелета. Длина второстепенных игл колеблется от 20 до 135 мкм. На основании соотношения размера игл и диаметра скелета устанавливается относительная длина игл: очень длинные, длинные, короткие и очень короткие. Соотношение размера игл и ширины их основания свидетельствует об относительной толщине второстепенных игл: очень толстая, толстая, тонкая, изящная.
Основные иглы закладываются на самом первом этапе становления скелета. Основой для начального этапа формирования первичной иглы служат первичные иглы, которые закладываются на продолжении лучей спикулы. Далее происходит постепенное нарастание скелета иглы от основания к дистальной части, причём скелетная ткань откладывается чётко по спирали.
Второстепенные иглы имеют не столь разнообразную форму, как основные иглы, и представлены, главным образом: 1) конусовидными иглами с округлым сечением основания 2)пирамидальными иглами с треугольным сечением основания. Второстепенные иглы не имеют углублений в основании игл и не образуют граней.
Симметрия скелета
Гомаксонный тип симметрии представляет собой сферические и производные от сферы скелеты с бесконечным числом одинаковых осей. Если число осей сокращено до 20 или 4-6, то уменьшается и число радиусов симметрии. Чаще всего такой скелет имеет форму, близкую к шару. Возможно, это оптимальное приспособление к планктонному образу жизни.
Монаксонный тип симметрии - число осей сокращено до одной, при этом скелет оказывается сплюснутым в направлении этой оси или, наоборот, вытянутым вдоль нее.
Гетерополярный тип симметрии - полюсы единственной оси не равнозначны. Число радиусов симметрии неопределенно, может равняться 3 или 2. В последнем случае образуются две половины скелета, которые можно теоретически совместить, повернув вокруг оси.
Билатеральная симметрия отражается не только на скелете, но и но в наружном расположении камер и сегментов. Этот тип симметрии предполагает дифференцирование скелета в направлении одной морфологической оси (условно верх-низ) и в направлении переднезадней оси, которая лежит в плоскости симметрии. Получающиеся половинки скелета – зеркальные.
Ось появляется при необходимости вращения, для сохранения равновесия и определения направление движения. Причем в случае, если направление движение безразлично, получается монаксонный тип, если же не безразлично, то гетерополярный или билатеральный. Такая геометрическая дифференциация вызывается планом биологической дифференциации (Мордухай-Болтовский, 1936).
В ряде случаев скелет радиолярий может быть асимметричен. Первичная асимметрия свойственна скелетным иглам с несколькими отростками у примитивных спумеллярий и насселярий. У последних на основе такого устройства скелета развивается асимметричный решетчатый панцирь с раструбом внизу. Вторичная асимметрия у высших представителей отряда феодарий развивается на основе радиальной симметрии.
Систематика и классификация
В настоящее время М. С. Афанасьевой и Э. О. Амоном разработана новая классификация, которая открывает возможность учитывать практически любые сведения об исследуемых организмах, и позволяет достаточно просто определить место в филогенетической схеме любого вновь рассматриваемого объекта изучения. Данная классификация разработана на уровне высоких и высших таксонов. В ней ранг Radiolaria повышен до типа.
Классификация М. С. Афанасьевой и Э. О. Амона (2006) включает 286 высоких таксонов радиолярий: 1 тип, 3 подтипа, 8 классов, 26 отрядов, 14 надсемейств, 130 семейств и 104 подсемейства, охватывающих 1140 родов радиолярий фанерозоя. Этой классификации мы и придерживаемся в данной работе.
Тип Radiolaria объединяет три подтипа: Polycystina Ehrenberg, 1838, Phaeodaria Haeckel, 1879, Collodaria Haeckel, 1879.
Размножение
Бесполое размножение может совершаться по типу вегетативного разделения на две части, после чего недостающие части достраиваются.
Половой процесс у радиолярий сопровождается образованием одно- или двужгутиковых одинаковых гамет, которые после копуляции формируются в зиготу.
У более сложных видов возможно формирование эмбриона.
Питание радиолярий
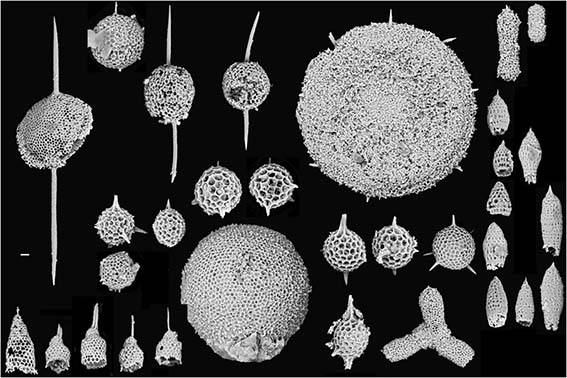
Образ жизни радиолярий
Радиолярии – исключительно морские организмы. Они живут в водах с нормальной солёностью – 32-38% и ведут планктонный образ жизни. Форма тела радиолярий приспособлена к парению в воде.
Радиолярии подразделяются на две группы по температуре обитания: относительно тепловодные формы и умеренно холодноводные. К первой группе относятся сфероидные и дискоидные формы. Ко второй относятся преимущественно циртоидные формы радиолярий. Наибольшая их распространённость наблюдается в экваториальном поясе, где наблюдается закономерное их уменьшение с глубиной. Небольшое число видов радиолярий приурочено к жизни только в холодных районах Мирового океана. То есть можно говорить об арктических и антарктических радиоляриях.
Среди радиолярий выделяют океанические и неритические виды, считающиеся единичными представителями.
Они отличаются способностью населять всю толщу вод океанов, начиная от поверхностных слоев и кончая самыми глубинными зонами (Основы палеонтологии, 1959).
Среди радиолярий выделяют как одиночные представители, так и колониальные. Они могут образовывать колонии от нескольких сантиметров до метра и более.
Радиолярии значение геологическое
Устанавливая и изучая историю развития условий обитания этих планктонных организмов, их географическое распространение в каждом веке фанерозоя, позволит установить географические ареалы отдельных видов, проследить их экологию, выстроить генетические цепочки, позволит создать стратиграфические шкалы на местном уровне и провести корреляцию этих разрезов.
Поделиться с друзьями