
Диатомовые водоросли представители
Латинское название Bacillariophyta
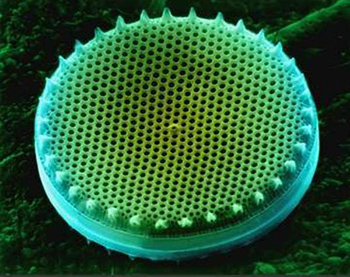
Диатомовые водоросли - это совершенно особая группа одноклеточных организмов, резко отличающаяся от остальных водорослей: клетка диатомовых снаружи окружена твердой кремнеземной оболочкой, называемой панцирем. Форма этого панциря настолько разнообразна, затейлива и причудлива, а структура его настолько тонка, изящна и красива, что его можно принять за творение искусного художника. Некоторые диатомеи красотой панциря могут поспорить с ювелирными изделиями изобретательного мастера.
Впервые о существовании диатомовых водорослей стало известно в начале XVIII в., когда в практику исследований были введены микроскопы Левенгука и сильно увеличивающая лупа.
Эта группа водорослей имеет несколько научных названий: диатомовые (Diatomeaea), кремнеземные (Kieselalgae) и бациллариевые (Bacillariophyta). Первое название они получили благодаря своему размножению делением панциря на две половинки, второе связано с наличием у клетки кремнеземной оболочки и, наконец, последнее название было дано по первому роду, научно описанному в 1788 г.,- бациллария, что обозначает «палочковидный». В русской литературе утвердилось название «диатомовые водоросли» или «диатомеи», наиболее современным научным латинским названием является Bacillariophyta.
Благодаря прозрачному панцирю и бурой окраске хлоропластов диатомовые легко различить под микроскопом среди остальных одноклеточных низших водорослей.
Диатомовые водоросли - одноклеточные микроскопические организмы, одиночно живущие или объединенные в колонии различного типа: цепочки, нити, ленты, звездочки, кустики или слизистые пленки. Колонии обычно микроскопические, но иногда макроскопические и тогда видны невооруженным глазом. Размеры клеток - от 4 до 1000 мкм, а у некоторых представителей - до 2000 мкм.
Исследованием клетки диатомей стали заниматься в начале прошлого столетия. При помощи светового микроскопа уже тогда удалось получить довольно полное и правильное представление о строении клетки и структуре панциря. Введение в практику альгологических исследований электронного микроскопа - трансмиссионного и сканирующего - за последние 10-15 лет позволило существенно дополнить наши знания о строении органелл клетки и тонкой структуре панциря, многие детали которой были неизвестны.
Строение Клетки Диатомовых Водорослей
Клетка диатомовых водорослей состоит из протопласта, окруженного кремнеземной оболочкой, называемой панцирем. Протопласт своим наружным уплотненным слоем (плазмалеммой) тесно примыкает к панцирю и заполняет его внутренние полости. Целлюлозной оболочки, имеющейся у большинства водорослей, здесь нет. Химический анализ панциря показал, что он состоит из аморфной формы кремнезема, напоминающей по составу опал, с плотностью 2,07. Толщина стенок панциря зависит от концентрации кремния в среде и колеблется в значительных пределах: у тонкостенных форм - от сотых до десятых долей микрометра, а у толстостенных достигает 1-3 мкм. Стенки панциря пронизаны мельчайшими отверстиями, обеспечивающими обмен веществ между протопластом и окружающей средой. Они снабжены также различными форменными элементами, которые составляют структуру панциря и служат основными таксономическими признаками при построении системы диатомей. Панцирь и его структура различимы уже при небольших увеличениях микроскопа. По форме панциря все диатомеи делятся на две группы: центрические - с радиально-симметричным панцирем и пеннатные - с двусторонне-симметричным панцирем.
Протопласт. Цитоплазма в клетках диатомей располагается постенным слоем или скапливается в центре клетки или у ее полюсов. Остальные участки клетки заполнены множеством вакуолей с клеточным соком, которые иногда сливаются в одну крупную вакуолю.
Ядро обычно шаровидное и располагается чаще всего близ центра клетки в цитоплазматическом мостике или в периферическом слое цитоплазмы. У некоторых диатомей оно имеет Н-образную форму. В ядре различают от 1 до 8 ядрышек.
Хлоропласты у диатомей довольно разнообразны по форме, величине и количеству в клетке. У большинства центрических диатомей они мелкие, многочисленные, в форме зерен или дисков, лишенные пиреноида. Реже они более крупные, по одному или несколько в клетке и имеют форму пластинки с ровными или изрезанными лопастными краями, с одним или несколькими пиреноидами. У пеннатных диатомей хлоропласты обычно крупные, пластинчатые, часто с лопастными краями или с перфорациями, они немногочисленные (один или два в клетке), занимают почти всю ее полость, как правило, с пиреноидами. Количество, величина и положение их различно даже среди представителей одного рода (табл. 10).
Окраска хлоропластов у диатомовых водорослей имеет различные оттенки желто-бурого цвета в зависимости от набора пигментов, среди которых преобладают бурые - каротин, ксантофилл и диатомин, маскирующие в живой клетке хлорофиллы а и с. После гибели клетки бурые пигменты растворяются в воде и зеленый хлорофилл становится ясно заметным.
Интенсивность окраски хлоропластов и их величина различны и зависят от образа жизни водорослей: у планктонных видов они золотисто-желтые, мелкие, дисковидные, а у донных и прикрепленных к субстрату - крупные пластинчатые, темно-бурые, поэтому большие скопления диатомей приобретают хорошо выраженную бурую или темно-бурую окраску.
В процессе фотосинтеза у диатомовых водорослей образуется масло в виде капелек различной величины, иногда в значительном количестве. Оно служит запасным питательным веществом, особенно в период, когда прекращается или задерживается деление клетки. Масло, экстрагированное из клеток диатомей, имеет запах рыбьего жира. Кроме масла, для некоторых видов характерно еще присутствие в клетках капель волютина, имеющих тусклый голубоватый блеск. Волютин нерастворим в эфире и при окраске живой клетки метиленовой синькой приобретает красновато-фиолетовый оттенок. Маленькие капли волютина распределяются по всей цитоплазме, а крупные (тельца Бючли) занимают определенное положение на концах клетки (виды семейства Nitzschiaceae) или по обеим сторонам центрального цитоплазматического мостика (роды семейства Naviculaceae). В качестве питательного вещества в клетках диатомей встречается еще лейкозин.
Панцирь и его строение. Панцирь диатомовых водорослей вырабатывается самой клеткой в процессе ее жизнедеятельности. Он состоит из двух почти равных частей и по конструкции напоминает коробку, закрытую крышкой. Наружная, большая часть панциря - эпитека, подобно крышке, находит своими краями на внутреннюю половину - гипотеку, соответствующую коробке (рис. 76). Эпитека и гипотека состоят из створки и пояскового ободка. Створку, принадлежащую эпитеке, называют эпивальвой, а гипотеке - гиповальвой. Створка имеет лицевую поверхность, плоскую или слегка выпуклую, и краевую загнутую часть, называемую загибом створки, иногда отличающуюся структурой. Загиб створки у одних диатомей низкий и выражен довольно слабо; у других он достаточно высокий и составляет значительную часть боковой поверхности панциря.
Створки бывают самых различных очертаний: круглые, эллиптические, яйцевидные, ромбические, ланцетные, треугольные, четырехугольные, булавовидные, серповидные, гитаровидные, клиновидные и т. д. Изменчивы и разнообразны концы створок: клювовидные, головчатые, оттянутые, тупые, острые и пр.
К каждой створке примыкает поясковый (соединительный) ободок, представляющий собой широкое или узкое кольцо, окаймляющее загиб створки, но не срастающееся с ним. Поясковый ободок эпитеки и своим свободным краем надвигается на поясковый ободок гипотеки и плотно его охватывает, с ним, однако, не срастаясь. Правда, у представителей некоторых родов поясковые ободки образуются только во время деления клетки, а эпивальва и гиповальва плотно соединяются друг с другом непосредственно краями загиба створки. Кроме того, у многих диатомей между загибом створки и поясковым ободком образуются еще вставочные ободки, от одного до многих. Каждый новый, более молодой вставочный ободок возникает всегда между загибом створки и предыдущим ободком. Они представляют собой дополнительные образования, отличающиеся не только формой, но и структурой. Форма вставочных ободков - один из характерных признаков рода. Они бывают воротничковидные, кольцевидные, полукольцевидные или состоят из отдельных сегментов, имеющих форму трапеции, ромба или чешуйки.
Наличие вставочных ободков в панцире имеет большое биологическое значение, так как они способствуют увеличению объема клетки и ее росту.
Часть панциря между эпивальвой и гиповальвой, т. е. поясковый ободок гипотеки и находящий на него поясковый ободок эпитеки, а если есть, то и вставочные ободки, называют пояском панциря.
Форма панциря зависит от очертаний створки. Он бывает шаровидный, палочковидный, седловидный, в виде апельсиновой дольки, низкого или высокого цилиндра, параллелепипеда или другой геометрической фигуры. Со стороны пояска он имеет обычно прямоугольную форму.
Характерной особенностью панциря является геометрическая правильность его строения, в связи с чем для представления о его форме очень важно учитывать соотношение осей и плоскостей симметрии.
При двусторонней симметрии у пеннатных диатомей определяют несколько осей и плоскостей симметрии, которые делят панцирь на симметричные половины. Известны три основные взаимно перпендикулярные оси симметрии: продольная, поперечная и центральная, проходящая через центр двух створок панциря.
Длина продольной оси определяет длину створки или панциря, длина поперечной оси - ее ширину, а длина центральной оси - высоту панциря. Кроме осей симметрии, различают три плоскости симметрии: продольную, проходящую вдоль панциря перпендикулярно к створкам и делящую его на две равные половины; поперечную, проходящую поперек панциря перпендикулярно к продольной плоскости и к створкам и не всегда делящую панцирь на две равные половины (если панцирь гетеропольный, а не изопольный, т. е. если концы створок неодинаковы); створковую, перпендикулярную к двум предыдущим, но параллельную створкам, т.е. проходящую через поясок панциря.
У центрических диатомей, обладающих радиальной симметрией, панцирь имеет только две оси и две плоскости симметрии. Одна ось - это диаметр створки, другая - центральная. Плоскость симметрии, проходящая через центр створки в любом направлении, всегда делит панцирь на две равные части; вторая плоскость симметрии - створковая, как и у пеннатных форм, идущая перпендикулярно к первой.
Форма створок и панциря в целом, а также соотношение осей и плоскостей симметрии имеют важное значение в систематике диатомовых водорослей. Однако главным признаком при построении их системы является структура кремнеземного панциря, которая представляет наибольшую трудность при изучении. Под структурой панциря, видимой в световой и электронный микроскопы, подразумевают наружный и внутренний его рисунки, специфические для разных таксонов. Структурные элементы на створках центрических диатомей имеют радиальное и тангенциальное расположение, а у пеннатных - двустороннее, или поперечное, расположение, т. е. их структура симметрична по отношению к продольной и поперечной осям. Реже у представителей некоторых родов панцири бывают асимметричными и не имеют ни одной плоскости симметрии, а иногда асимметричность выражается только в структуре створок.
<
Главная особенность стенок панциря состоит в том, что они пронизаны регулярно повторяющимися мельчайшими отверстиями - ареолами, обычно затянутыми снаружи или внутри тонкой перфорированной пленкой, получившей латинское название "велум". Отверстия в стенке панциря необходимы для сообщения протопласта клетки с окружающей средой. При изучении диатомовых водорослей в световом микроскопе казалось, что у некоторых видов бесструктурный панцирь, и только внедрение в практику альгологических исследований электронного микроскопа показало, что и эти панцири также имеют чрезвычайно тонкую пористую стенку. Отверстия, пронизывающие створку, занимают обычно 10-75 % ее площади, а характер расположения этих отверстий и их количество специфичны для разных родов и видов. Но на створках есть и участки, лишенные отверстий, например центральное, осевое и боковые поля у шва (см. ниже) и некоторые скульптурные детали структуры. У центрических диатомей ареолы располагаются радиально и тангенциально; у пеннатных - поперечными рядами, либо параллельными друг другу, либо к краям створки слегка расходящимися (радиальные ряды) или, наоборот, сходящимися (конвергентные ряды).
Иногда ареолы располагаются так, что, кроме поперечных, образуются еще и продольные или взаимно перекрещивающиеся косые ряды.
Примечательная особенность пеннатных диатомовых водорослей - наличие осевого поля, представляющего собой бесструктурную узкую или широкую полосу по продольной оси створки. У некоторых диатомей осевое поле расширяется на середине створки, образуя среднее поле, которое бывает круглым, ромбическим, четырехугольным, иногда доходящим до краев створки.
Большинство диатомей пеннатного типа характеризуется еще одним признаком - присутствием шва, представляющего собой короткую или длинную щель или две щели (ветви шва), прорезывающие стенку створки и идущие вдоль створки от ее концов к середине. Строение шва весьма различно - от простого щелевидного до так называемого каналовидного. Примитивный щелевидный шов представлен двумя короткими изолированными щелями, не доходящими до середины створки. У представителей некоторых родов такой шов находится всего на одной створке, иногда только у одного из ее концов, у других - на обеих створках. Хорошо развитый щелевидный шов, характерный для водорослей из семейства навикуловых (Naviculaceae), представлен двумя длинными щелями, или ветвями, шва, проходящими по обеим створкам и соединяющимися на середине каждой створки центральным узелком, а у концов створки заканчивающимися конечными, или полярными, узелками (рис. 83). Щели шва в толще створки коленчато-изогнуты, так что в поперечном разрезе они имеют вид лежащей буквы V (рис. 84). Щель шва, открывающуюся внутрь клетки, называют внутренней, а открывающуюся наружу - наружной. В центральном узелке обе ветви шва соединяются друг с другом, заканчиваясь здесь центральной порой, а у концов створки - конечной порой.
Центральный узелок представляет собой внутреннее утолщение стенки створки, выпуклость на ее внутренней поверхности, а конечные узелки - внутреннее и наружное утолщения стенки створки.
Наиболее сложное устройство имеет так называемый каналовидный шов - канал, расположенный в складке стенки створки. С внешней средой он сообщается узкой щелью, а в полость клетки открывается рядом отверстий с кремнеземными перегородками - фибулами. Каналовидный шов присущ водорослям семейств эпитемиевых (Epithemiaceae), нитцшиевых (Nitzschiaceae) и сурирелловых (Surirellaceae). Он также имеет центральный узелок, но его положение на створке различно у представителей разных родов. У водорослей из рода эпитемия (Epithemia) ветви каналовидного шва соединяются под углом и приближены к брюшному краю; у видов рода ропалодия (Rhopalodia) шов тянется по спинному краю; у представителей рода нитцшия (Nitzschia) находится в киле, расположенном вдоль одного из краев створки, а у видов рода дентикула (Denticula) проходит более или менее эксцентрично к продольной оси створки. У водорослей из родов сурирелла (Surirella) и кампилодискус (Campylodiscus) каналовидный шов лежит на краю крыла створки, находящегося на границе с загибом створки и опоясывающего ее. Поэтому при рассмотрении панциря со стороны створки он не виден. И только у представителей рода цилиндротека (Cylindrotheca) шов спирально окружает панцирь.
Биологическое значение шва в жизни диатомей очень велико: помимо сообщения протопласта клетки с внешней средой, с помощью шва клетки довольно быстро передвигаются по субстрату и в толще воды. В филогенетическом отношении возникновение шва является прогрессивным признаком; он присущ более молодым видам, которые в современных морях и океанах составляют свыше 70 % от общего числа диатомей.
Кроме названных структур, у большинства диатомовых водорослей на внешней и внутренней поверхности створок имеются различные образования в виде полых или сплошных выростов, выпуклостей, рогов, щетинок, шипов, шипиков, бороздок, камер, ребер и пр., которые выполняют определенные функции: выделяют слизь, объединяют клетки в колонии, увеличивают поверхность панциря у планктонных видов, обеспечивая плавучесть клетки в воде.
У некоторых диатомей, имеющих вытянутые створки, на внутренней поверхности вставочных ободков образуются кремнеземные перегородки, или септы, вдающиеся в полость панциря параллельно плоскости створок. Септы возникают или по всей внутренней поверхности вставочного ободка, или только в одном из его концов. Они обычно хорошо видны со стороны пояска, различны по положению, форме и размерам и имеют одно или несколько отверстий. У диатомей с гетеропольным панцирем септы чаще всего возникают только в широком его конце (род Licmophora), с изопольным панцирем - в любом или в обоих его концах (роды Tetracyclus, Tabellaria). Септы могут быть узкими, или они глубоко вдаются в полость панциря, вплоть до его середины.
У небольшого числа диатомей образуется еще один тип перегородок, так называемые псевдосепты, развивающиеся на внутренней стороне самой створки и вдающиеся в полость панциря в виде короткой и довольно грубой перегородки, видимой со створки и с пояска. В отличие от септ псевдосепты всегда перпендикулярны к створке и возникают одновременно с ней. Представителям рода мастоглойя (Mastogloia) свойственны особые образования - камеры, представляющие многоугольные, реже удлиненные пустоты в стенке панциря, открытые внутрь клетки или наружу круглыми отверстиями.
Некоторые детали структуры бывают не видны в световом микроскопе, а обнаруживаются только при больших увеличениях с помощью электронного микроскопа. Все перечисленные структуры обладают четкой, правильной формой и определенным количеством элементов на единицу поверхности. Большинство из них выполняет определенную функцию, обеспечивая приспособляемость диатомей к условиям существования.
Диатомовые водоросли способы питания
Диатомовые водоросли преимущественно фотоавтотрофные организмы, которые в процессе фотосинтеза образуют органическое вещество. В хлоропластах диатомей обнаружено девять пигментов: хлорофиллы a и c, - и -каротины и пять ксантофиллов - фукоксантин, диатоксантин, неофукоксантины A и B и диадиноксантин. Состав и количество пигментов непостоянно и зависит от интенсивности света, его качества, содержания в воде биогенов, а также от возраста клетки и особенностей ее жизнедеятельности. Количество хлорофилла уменьшается в старых клетках, а недостаток азота и фосфора резко снижает содержание хлорофилла a. Недостаток питательных веществ в воде даже при высокой интенсивности света ведет к уменьшению количества пигментов, а обилие питательных веществ даже при слабой освещенности способствует их образованию. Конечным продуктом фотосинтеза являются жиры, а не углеводы.
Интенсивность фотосинтеза на единицу биомассы у планктонных и бентосных диатомей неодинаковая. У бентосных форм она значительно выше, так как их хлоропласты крупнее и имеют более интенсивную окраску. Кроме того, у подвижных форм фотосинтез протекает активнее, чем у неподвижных, и значительно усиливается в период деления клеток. Условия для фотосинтеза у поверхности воды довольно близки к условиям воздушной среды, но резко меняются с погружением водорослей на глубину.
Планктонные диатомовые водоросли, обитающие в пелагиали морей, могут существовать на глубине 100 м и более при большой прозрачности воды. Однако с глубиной меняется не только интенсивность освещения, но и качество света вследствие различного поглощения лучей солнечного спектра разной длины волны, что по-разному отражается на разных видах.
Среди планктонных и бентосных диатомей существуют светолюбивые и тенелюбивые виды, отличающиеся интенсивностью фотосинтеза и коэффициентом использования солнечной энергии при одинаковой радиации. У светолюбивых видов максимум фотосинтеза приходится на полдень, а у тенелюбивых - на утренние и послеполуденные часы.
Изучение диатомовых водорослей в культурах позволило выявить большую пластичность диатомей в усвоении как минеральных, так и органических веществ.
Особую роль в жизни диатомовых водорослей играет кремний, который им необходим для построения панциря. Усвоение его происходит в соответствии с ритмом деления клеток и зависит от химических и физических свойств окружающей среды. Деление клеток диатомей происходит нормально, если в воде находится не менее 5 мг/л кремния, а при содержании его около 0,5 мг/л деление прекращается.
Усваивается кремний диатомовыми водорослями в виде кремниевой кислоты и органических соединений кремния. Потребность в кремнии у диатомей различная и зависит от местообитания и физиологического состояния клеток. Так, например, бентосные виды, имеющие толстостенный панцирь, нуждаются в большем количестве кремния по сравнению с планктонными формами, обладающими тонкостенным панцирем. В период обильного размножения, которое происходит обычно весной, а у некоторых видов и осенью, диатомеи испытывают наибольшую потребность в кремнии: недостаточное содержание его в воде вызывает замедление темпов деления и приводит к уменьшению толщины панциря.
Помимо неорганических веществ, диатомовым водорослям для роста и развития также нужны небольшие количества органических веществ. Очень хорошее воздействие на них оказывает витамин B . Изучение органического питания диатомей показало, что они нуждаются в витаминах группы В больше, чем другие водоросли.
Некоторые диатомовые водоросли вообще могут переходить от автотрофного питания к гетеротрофному. Известны даже формы с бесцветными хлоропластами или вовсе без них - эти водоросли являются уже облигатными гетеротрофами.
Размножение Диатомовых Водорослей
Деление. Чаще всего диатомеи размножаются вегетативным делением клетки на две половины; этот процесс обычно происходит ночью или на рассвете. Темпы деления различны у разных видов и могут меняться даже у одного вида в зависимости от сезона или условий окружающей среды. Весной и в начале лета наблюдается максимальное развитие диатомовых в результате их интенсивного деления. Наличие в воде биогенных веществ способствует делению и росту диатомей.
Опыты показали, что в культуральной среде некоторые планктонные виды могут делиться до 3-8 раз в сутки. Бентосные виды делятся гораздо реже - один раз в 4 дня. Известны случаи еще более редкого деления - один раз за 25 дней. Но эти сведения не абсолютны, и темпы деления могут меняться в зависимости от широтного расположения водоема, его физико-химического режима и, конечно, от особенностей вида.
Процесс деления клетки у диатомей своеобразен из-за наличия твердого панциря. Сначала в протопласте начинают скапливаться капельки масла, а сам протопласт значительно увеличивается в объеме, вследствие чего эпитека и гипотека панциря расходятся, оставаясь соединенными только краями своих поясковых ободков. Протопласт делится на две равные части, а вместе с ним и хлоропласты. Если хлоропласт один, то он делится пополам; если их много, то в дочерние клетки сначала попадает половина их, а затем они делятся, в результате в дочерних клетках образуется исходное количество хлоропластов. Ядро делится митотически, часто с хорошо заметными хромосомами и центросомой на каждом из образовавшихся полюсов. После окончательного разделения клетки на две каждая из дочерних клеток, получившая лишь половину материнского панциря, сразу же "достраивает" недостающую половину, но обязательно внутреннюю, т.е. гипотеку. Дополнительные образования панциря - вставочные ободки, септы и другие структурные элементы - возникают вскоре после сформирования новой гипотеки. Таким образом, возникшие в результате деления две дочерние клетки оказываются по размерам несходными: одна клетка, получившая эпитеку, сохраняет размеры материнской клетки, а другая, получившая материнскую гипотеку, ставшую в новой клетке эпитекой, приобретает меньшие размеры. В результате после многократных делений происходит постепенное уменьшение размеров клеток у половины каждой данной популяции: у центрических диатомей уменьшается диаметр клеток, а у пеннатных - длина и отчасти ширина клеток. Установлено, что у некоторых видов в процессе делений размеры клеток уменьшаются почти в 3 раза по сравнению с первоначальными.
Микроспоры. У многих планктонных диатомей обнаружены так называемые микроспоры - мелкие тельца, возникающие в клетках в количестве от 8 до 16 и более, а у некоторых видов их бывает и более 100. Наблюдались микроспоры со жгутиками и без них, с хлоропластами и бесцветные. Наиболее часто микроспоры развиваются у видов рода хетоцерос (Chaetoceros), и наблюдалось даже их прорастание (рис. 90).
Процесс образования микроспор цитологически не изучен, и природа их точно не установлена.
Половой процесс и образование ауксоспор. Ауксоспорами, т.е. "растущими спорами", называют такие, которые при своем образовании сильно разрастаются и затем прорастают в клетки, резко отличающиеся по размерам от исходных. Способность образовывать ауксоспоры свойственна только диатомовым водорослям, но до сих пор не удалось еще полностью объяснить этот процесс и порождающие его причины. Образование ауксоспор, по всей вероятности, вызывается различными причинами. Согласно наиболее распространенному мнению, оно наступает вследствие многократных делений, приводящих, как это описано выше, к мельканию клеток. Достигнув минимальных размеров, клетки развивают ауксоспоры, что приводит к восстановлению их размеров. Однако другие исследователи считают, что ауксоспорообразование связано просто со старением клеток, так как его нередко удавалось наблюдать и тогда, когда клетки еще не достигли своих минимальных размеров. С этих позиций ауксоспорообразование рассматривается как процесс "омоложения" клетки. Кроме того, есть наблюдения, свидетельствующие о развитии ауксоспор при изменении условий окружающей среды, например при резком понижении температуры воздуха или воды.
Каковы бы ни были причины, способствующие возникновению ауксоспор, установлено главное: ауксоспорообразование всегда связано с половым процессом. У диатомовых водорослей встречаются все три типа полового процесса, вообще известные у водорослей, - изогамный, анизогамный и оогамный, а также некоторые формы редуцированного полового процесса (рис. 91). У пеннатных диатомей половой процесс во всех случаях состоит в сближении двух клеток, в каждой из которых створки раздвигаются и происходит редукционное деление ядра, после чего гаплоидные ядра попарно сливаются и образуется одна или две ауксоспоры. У центрических диатомей по парное сближение клеток отсутствует и ауксоспора образуется из одной клетки, в которой сначала происходит деление материнского диплоидного ядра на четыре гаплоидных ядра, два из них затем редуцируются, а два сливаются в одно диплоидное ядро и образуется ауксоспора.
Все диатомовые водоросли - диплоидные организмы, а гаплоидная фаза у них бывает только перед слиянием ядер в ауксоспоре. Как в первом, так и во втором случае после слияния ядер образуется зигота, которая сразу, без стадии покоя, резко увеличивается в размерах и развивает ауксоспору. По положению и связи с материнской клеткой ауксоспоры бывают разных типов: свободная ауксоспора, конечная, боковая, интеркалярная и полуинтеркалярная.
После созревания ауксоспоры в ней развивается новая клетка, у которой сначала образуется эпитека, а затем гипотека. Первую клетку, возникшую из ауксоспоры, называют инициальной. По размерам она значительно превышает исходную.
Покоящиеся споры. Образованию покоящихся спор обычно предшествует или обильная вегетация вида, или наступление неблагоприятных условий. Протопласт клетки сжимается, округляется, и на его поверхности появляется сначала первичная створка споры, а затем вторичная, обе плотно соединяются краями (поясок у них отсутствует). Створки часто отличаются структурными элементами, они покрыты шипиками, выростами и некоторыми другими образованиями. Обычно у диатомовых водорослей в клетке развивается только одна спора. Спустя определенное время покоящаяся спора, подобно ауксоспоре, увеличивается в объеме и дает начало новой клетке, вдвое большей по сравнению с исходной.
Покоящиеся споры обычно образуют многие морские неритовые диатомеи, а также некоторые пресноводные виды. У представителей многих родов они возникают периодически как обычное явление в жизненном цикле.
Биологические особенности диатомовых водорослей