
Класс Ресничные черви или турбеллярии
Turbullaria
Тип Плоские черви - Plathelminthes
Общая характеристика
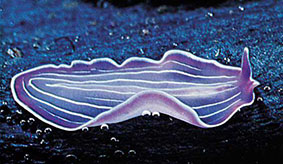
Ресничные черви, или турбеллярии, живут в морях, океанах и пресных водах. Большинство турбеллярий ведет донный (бентосный) образ жизни. Обычно они ползают по дну или по водным растениям. Среди турбеллярий есть и такие, которые живут в почве и на ее поверхности — под листвой и т. п. Многие турбеллярии — хищники, и лишь немногие приспособились к паразитическому образу жизни. Турбеллярий насчитывается около 3000 видов.

Внешнее строение ресничных червей
Размеры ресничных червей колеблются чаще всего в пределах нескольких миллиметров, реже сантиметров, среди них очень много мелких форм, размеры которых не превышают 1—2 мм. Однако среди турбеллярий имеются и более крупные черви. Так, байкальский червь Polycotylus достигает 30 см, а некоторые наземные тропические формы имеют длину 50—60 см.
Тело турбеллярий в большинстве случаев сплющено в дорзовентральном направлении, листовидной формы, одна-ко среди мелких видов некоторые имеют более пли менее веретенообразную форму.
У большинства турбеллярий на теле нет никаких придатков. Только у некоторых на головном конце имеются два выроста в виде небольших щупалец. Движения турбеллярий разнообразны. Они обусловлены, с одной стороны, движением ресничек, покрывающих тело турбеллярий, а с другой — сокращением мускулатуры.
Кожно-мускульный мешок
Поверхность тела ресничных червей покрыта однослойным ресничным мерцательным эпителием. Под ним расположены многочисленные одноклеточные (реже многоклеточные) слизистые, клейкие и белковые железы, протоки которых открываются наружу среди эпителиальных клеток. Слизистые железы выделяют слизь, облегчающую скольжение турбеллярий. Выделения клейких желез застывают в виде нитей, на которых животные могут временно подвешиваться на поверхностной пленке воды или подводных предметах. Белковые железы образуют ядовитый секрет, имеющий защитное значение.
Многие эпителиальные клетки содержат так называемые рабдиты. Это сильно преломляющие свет палочки, расположенные внутри клеток. Они представляют собой «оформленный секрет» клеток. Рабдиты формируются непосредственно в эпителиальных клетках или в клетках, расположенных глубже,— в паренхиме. Последние связаны с эпителиальными клетками цитоплазматическими мостиками, по которым рабдиты продвигаются к поверхности.
При малейшем раздражении рабдиты выбрасываются из клеток наружу и расплываются в слизистую массу. Они состоят из ядовитых веществ и представляют собой средство защиты и нападения. Во всяком случае, известно, что многие ресничные черви несъедобны для других животных.
Под кожным эпителием, отделенные от него тонкой базальной перепонкой, расположены слоями мускульные волокна. Непосредственно под эпителием расположен сплошной кольцевой, или поперечный, слой мышечных волокон. Этот слой называется так потому, что оси мускульных клеток расположены поперек оси тела червя. Сокращение этих мышц обусловливает сжатие тела. Под кольцевым слоем обычно расположен слой так называемых косых, или диагональных, мышц. Оси мышечных волокон, составляющих этот слой, расположены перпендикулярно друг к другу и под углом к кольцевому слою. Наконец, третий слой состоит из мышечных волокон, вытянутых вдоль тела животного. Это слой продольных мышечных волокон. Все мышечные слои состоят из гладких мышечных волокон. Мышцы вместе с кожным эпителием образуют кожно-мускульный мешок, очень характерный не только для плоских, но и для других типов червей, хотя число мышечных слоев и их последовательность могут быть различны.
Кроме мышц, входящих в состав кожно-мускульного мешка, у них имеются еще пучки мышц, тянущихся от спинной части кожно-мускульного мешка к брюшной. Это — дорзовентральные мышечные пучки. Совокупность всей описанной мускулатуры обусловливает все довольно сложные движения тела турбеллярий.
Паренхима
Как уже было отмечено, внутри кожно-мускульного мешка все пространство между различными органами заполнено паренхимой, состоящей в основном из рыхло расположенных клеток неопределенной формы; часто эти клетки снабжены отростками, с межклеточным веществом между ними.
Паренхима представляет собой рыхлую соединительную ткань мезодермического происхождения. Среди основных клеток паренхимы расположены многочисленные мускульные волокна, железистые, рабдитные клетки и т. п.
Пищеварительная система
У турбеллярий, как у кишечнополостных и гребневиков, Пищеварительная система замкнутая, т. е. рот является единственным отверстием, через которое поглощается нища и выбрасываются её "непереваренные остатки — экскременты. Для большинства турбеллярий характерно разделение кишечника на два отдела: переднюю эктодермическую глотку и среднюю слепозамкнутую энтодермическую кишку. Рот всегда помещается на брюшной стороне, но может быть ближе к переднему или заднему концу, а иногда может располагаться в центре брюшной поверхности.
У некоторых турбеллярий глотка может отсутствовать или иметь форму короткой простой трубки, средняя кишка у них совсем отсутствует, а пищеварительные клетки расположены в паренхиме, не образуя пищеварительной полости. Такое весьма простое строение органов пищеварения свойственно низшим турбелляриям, преимущественно обитающим в морях и объединяемым в отряд бескишечных (Acoela).
У всех прочих ресничных червей (турбеллярий) глотка хорошо развита, причем чаще всего она представляет собой трубку с очень мускулистыми стенками, помещающуюся в особом влагалище, из которого глотка может высовываться наружу. Такая глотка представляет собой ловчий или сосательный аппарат.
Средняя кишка может иметь различное строение. У одних турбеллярий глотка ведет в среднюю кишку мешковидной формы, не имеющую никаких разветвлений. Это имеет место у мелких турбеллярий.
У крупных турбеллярий кишечник более или менее сильно разветвлен, от мешковидной части кишки отходят ветви: одна вперед к голове и много парных ветвей, идущих во все стороны. Эти выросты кишечника, в свою очередь, разветвляются. Такое строение кишечника наблюдается у морских турбеллярий, относящихся к отряду многоветви-стокишечных (Polycladida). Разветвленность средней кишки и радиальное расположение ее ветвей у многоветвистокишечных дало повод сравнивать среднюю кишку этих турбеллярий с гастроваскулярной системой кишечно-полостных.
Наконец, у турбеллярий из подотряда трехветвистокишечных (Tricladida) главной кишки нет и прямо от глотки отходят три ветви средней кишки. Одна ветвь идет вперед к головному отделу, а две направлены к заднему концу тела. Все эти ветви кишечника, в свою очередь, разветвляются. К этому подотряду относятся многие пресноводные турбеллярий.
Степень разветвленности кишечника различных турбеллярий, несомненно, связана с размерами животных. Не считая бескишечных, наиболее мелкими среди турбеллярий будут формы с не разветвленным кишечником.
Наибольшей степени разветвленности кишечник достигает у более крупных — многоветвистокишечных и трехветвистокишечных турбеллярий. Это объясняется отсутствием у турбеллярий кровеносной системы. Средняя кишка не только является органом пищеварения, но и выполняет функцию распределения пищи по всему телу, подобно гастроваскулярной системе медуз и гребневиков. Стенки средней кишки выстланы однослойным эпителием, состоящим из клеток с закругленными расширенными концами, среди которых расположены особые железистые клетки. Эти клетки выделяют в полость кишки пищеварительные ферменты. Однако переваривание пищи в полости кишки происходит только частично. Мелкие частицы пищи захватываются клетками эпителия кишечника и перевариваются внутри этих клеток.
Таким образом, в отношении процесса пищеварения турбеллярий мало отличаются от кишечнополостных. Клетки средней кишки обладают фагоцитарной функцией, и пищеварение у турбеллярий также, в значительной степени, внутриклеточное.
Анального отверстия и задней кишки у турбеллярий, как и у всех плоских червей, нет. Однако у некоторых турбеллярий имеются особые поры, при помощи которых полость кишечника сообщается с внешней средой. Значение этих пор не выяснено.
Выделительная система
Органы выделения впервые возникают у ресничных червей. Они представлены системой сильно разветвленных каналов, часто образующих перемычки, или анастомозы. Наиболее тонкие канальцы замыкаются слепо концевыми, или терминальными, клетками, а главные каналы открываются выделительными отверстиями. Терминальные клетки грушевидной формы, часто со звездчатыми отростками, расположены они непосредственно в паренхиме. Внутри клеток имеется полость, в которой помещается пучок длинных ресничек. Пучок ресничек находится в непрерывном колебательном движении, напоминающем колебания пламени свечи, за что эти клетки называют пламенными клетками. Полость терминальной клетки продолжается в ее отростке. Это и есть начало выделительного канала. Далее к отростку клетки примыкает ряд удлиненных клеток, сквозь которые проходит канал. Канальцы, отходящие от близко расположенных пламенных клеток, соединяются в более крупные протоки, затем эти протоки впадают в еще более крупные, открывающиеся одним или несколькими отверстиями наружу.
Описанные органы выделяют из тела избыток воды, а также жидкие продукты диссимиляции. Продукты распада органических веществ диффузно проникают из паренхимы в полость выделительной клетки и движением мерцательного пламени гонятся по каналам, которые также выстланы ресничками, и, наконец, выделяются наружу.
<
Важнейшей чертой выделительных органов ресничных червей (и всех плоских червей) является наличие особых терминальных клеток, замыкающих выделительные каналы. Такого типа выделительные органы беспозвоночных называют протонефридиями.
У различных турбеллярий выделительные органы развиты неодинаково. Они менее развиты у морских форм (многоветвистокишечные и бескишечные турбеллярии), вероятно, потому, что в условиях жизни в соленых водах организм не перегружается водой.
Нервная система
У некоторых наиболее примитивных ресничных червей из отряда бескишечных нервная система представляет собой диффузное нервное сплетение, более густое на переднем конце тела помещается скопление нервных клеток, образующих зачаточный головной ганглий, от которого отходят нервные стволы, идущие почти радиально.
У многоветвистокишечных ресничных червей мозговой ганглий расположен близко от центра тела (у округлых форм) или сдвинут к переднему концу (у вытянутых в длину форм). От него радиально расходится до 11 пар нервных стволов, связанных поперечными перемычками, или комиссурами. Обычно наиболее развита задняя пара нервных стволов. В результате образуется довольно правильная нервная сеть, особенно ясно выраженная у форм с центрально-расположенным нервным ганглием.
Органы чувств ресничных червей, глаза
Органы чувств представлены прежде всего осязательными клетками, особенно многочисленными на переднем конце и по бокам тела. Имеющиеся у некоторых ресничных червей или турбеллярий головные щупальца служат органами химического чувства.
У многих турбеллярий (бескишечных, некоторых Catenulida, Seriata и др.) в тесной связи с головным ганглием расположен статоцист, в виде замкнутого пузырька со статолитом внутри. Статоцист орган ориентации животного в пространстве. При изменении положения тела червя сигнал от статоциста передается через нервную систему мускулатуре турбеллярий до тех пор, пока последняя не примет нормального положения.
У большинства турбеллярий имеется одна или несколько пар глаз (у некоторых наземных планарий более 1000) иного строения, чем уже известные нам глаза медуз. Глаза помещаются непосредственно под кожным эпителием и состоят из пигментного бокала и зрительных клеток. Пигментный бокал, чаще состоящий из одной гигантской клетки, имеет форму чаши, обращенной вогнутой частью к периферии. Клетка (или клетки, если бокал многоклеточный) наполнена пигментом, и ядро помещается в ее выпуклой части. В пигментный бокал погружена одна или несколько зрительпых клеток своеобразной, булавовидной формы. Расширенные концы этих клеток заканчиваются светочувствительными палочками или колбочками. Изогнутые части зрительных клеток обращены к поверхности тела, и к ним подходят нервы головного ганглия. Благодаря такому расположению клеток лучи света сначала проходят через плазму зрительной клетки, а затем уже попадают на светочувствительную часть клетки. (У других животных светочувствительная часть клетки обращена непосредственно к свету). Поэтому глаза такого строения, как у турбеллярий, называются обращенными или инвертированными.
Размножение
Подавляющее большинство ресничных червей — гермафродиты. Половые органы у ресничных червей устроены крайне сложно и разнообразно в разных группах. Они отличаются по числу гонад, их структуре, присутствию многих дополнительных образований половой системы. Так, мужские половые железы — семенники — могут быть крупными одиночными или парными либо мелкими многочисленными образованиями. Женские половые железы — яичники — обычно парные, но могут быть и одиночными, и многочисленными. У более примитивных турбеллярий яичники простые. В них формируются яйца, которые содержат некоторое количество желтка, а также скорлупкового вещества. Такие яйца называют энтолецитальными. У более высокоорганизованных турбеллярий яичники дифференцируются на отделы: один из них, большой, продуцирует только питательные желточные клетки, а другой, маленький,— яйцеклетки. Эти отделы могут превращаться в самостоятельные парные органы: собственно яичники и желточники. Образующиеся яйцеклетки совсем лишены желтка. После оплодотворения их окружают желточные клетки, а затем вокруг них формируется общая оболочка. Такие яйца называют эктолецитальными.
Протоки половых желез — семяпроводы и яйцеводы — обычно парные, в нижнем отделе сливаются в непарные образования. Они могут открываться самостоятельно мужским и женским половыми отверстиями на брюшной стороне тела или в общую половую клоаку.
Низшие турбеллярий лишены женских выводных протоков. Так, у некоторых бескишечных ресничных червей яйцеводы отсутствуют. Сперма вносится партнером, который прорывает покровы червя совокупительным органом. Сперма попадает в паренхиму и оплодотворяет находящиеся там яйца. Откладка яиц возможна через разрыв стенок тела или через рот, как у кишечнополостных.
Сложное строение гермафродитной половой системы ресничных червей разберем на примере обычной в пресных водах молочной планарии (Dendrocoelum lacteum).
Мужские половые органы состоят из многочисленных мелких семенников, расположенных в паренхиме по бокам всего тела. От семенников отходят тончайшие семявыносящие канальцы, которые впадают в два семяпровода, направляющихся назад. Позади глотки семяпроводы впадают в семенной мешок. В задней части семенной мешок переходит в совокупительный орган, пронизанный семяизвергательным каналом. Совокупительный орган при совокуплении выдвигается через половую клоаку и вводится в половое отверстие другой особи.
Женский половой аппарат состоит чаще всего из одной пары яичников, расположенных в передней части тела. От яичников отходят два длинных яйцевода, направляющихся по бокам тела назад и сливающихся в непарный яйцевод, который открывается в половую клоаку рядом с карманом совокупительного органа.
На всем протяжении парных яйцеводов в них открываются протоки многочисленных желточников, в которых образуются особые желточные клетки, богатые питательными материалами.
В половую клоаку открываются еще два органа: копулятивная сумка — складчатый мешок с довольно тонким стебельчатым каналом и мускулистый железистый орган. Его значение не выяснено.
При спаривании молочных планарий совокупительный орган вводится в половое отверстие и через половую клоаку в копулятивную сумку другой особи. Таким образом, сперма прежде всего попадает в копулятивную сумку, а из нее — в яйцеводы, в ту их часть, которая расположена вблизи яичников. Оплодотворение происходит при выходе яиц из яичника в яйцевод. Затем яйца, продвигаясь по яйцеводам мимо отверстий желточных протоков, окружаются желточными клетками и попадают в половую клоаку. Здесь вокруг яиц вместе с желточными клетками образуется кокон из выделений желточных клеток и особых скорлуповых желез. Отложенный кокон подвешивается к подводным предметам.
Развитие
У ресничных червей с энтолецитальными яйцами происходит полное неравномерное дробление по спиральному типу, напоминающее дробление яиц кольчатых червей, немертин и моллюсков.
Развитие турбеллярий обычно прямое, лишь у некоторых групп наблюдается метаморфоз. У морских многоветвистокишечных ресничных червей из яйца выходит своеобразная мюллеровская личинка яйцевидной формы. Вначале она обнаруживает черты радиальной симметрии, а затем все более приобретает билатеральную симметрию. Впереди рта, расположенного на брюшной стороне, имеется 8 лопастных выростов, покрытых ресничками. Такая личинка ведет планктонный образ жизни, и этим обеспечивается расселение морских турбеллярий. Личинки морских турбеллярий переносятся морскими течениями на большие расстояния и постепенно превращаются во взрослых животных. При этом их рот сдвигается вперед, околоротовые лопасти уменьшаются, а все тело сплющивается. Личинка опускается на дно и окончательно приобретает билатеральную симметрию.
Развитие эктолецитальных яиц проходит по-иному. У описанной выше молочной планарии в коконе содержится от 20 до 40 яиц и около 80— 90 тысяч желточных клеток. Последние окружают каждую яйцеклетку, а позднее сливаются и образуют синцитий. Бластомеры оказываются разобщенными и погруженными в общую массу желтка. Они образуют три группы клеток, две из которых обеспечивают поглощение желтка зародышем, а из третьей формируется сам зародыш. Развитие прямое: из кокона отрождаются маленькие планарии.
Бесполое размножение наблюдается у некоторых турбеллярий из отрядов Macrostomida, Catenulida и Seriata (подотряд Tricladida). Оно заключается в поперечном делении червей. У некоторых форм, например у Microstomum lineare, бесполое размножение происходит все лето и лишь осенью сменяется половым. При бесполом размножении посередине тела появляется перетяжка, и у задней половины начинается образование рта и глотки. Задолго до разделения червя надвое дочерние особи также начинают делиться и появляются перетяжки II, III и т. п. порядков. Так образуется цепочка из делящихся зооидов.
Классификация >> подробнее>>
Раньше классификация ресничных червей основывалась в основном на строении их кишечника (его разветвленности), которое, как указывалось выше, в значительной степени зависит от размеров тела животных. В настоящее время при выделении главных систематических категорий это класса исходят из комплекса признаков: строения половой системы (преимущественно женских гонад), типа яиц, характера дробления, строения выделительной системы. Класс турбеллярий разделяется на два подкласса. 1. Архоофоры (Archoophora); 2. Неоофоры (Neoophora).
Поделиться с друзьями
