Морские гидроидные полипы
Leptolida
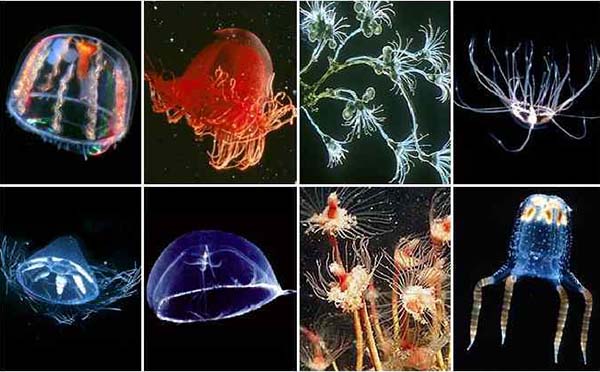
Некоторые морские гидроидные полипы одиночны, как гидры, но в подавляющем большинстве колониальны. Колонии разрастаются путем почкования большого количества полипов, называемых гидрантами, не отделяющихся от колонии. Часто колонии образуют стелющиеся по субстрату стволы, от которых отходят ветви, несущие гидрантов. Колонии полипов могут состоять из очень большого количества особей. Морские полипы отличаются от пресноводных гидр тем, что, помимо полипов, или гидрантов — бесполых особей, они образуют также путем почкования особые половые особи — медузы.
Строение гидроидных полипов
Отдельные гидранты обычно небольших размеров и по своему строению очень похожи на гидр. Их важнейшие отличительные особенности следующие. Часть тела, окружающая рот, у некоторых видов образует хоботок. Щупальца изнутри заполнены тяжем энтодермальных клеток и не имеют полости. Расположение и число щупалец может быть различно. Так, у гидроида Corype булавовидные щупальца не собраны в венчик вокруг рта, а разбросаны по всему телу. У другого гидроида — Pennaria — имеются два вида щупалец, различно расположенных, а у Obelia и многих других имеется один околоротовой венчик щупалец.
Веточки колонии, на которых сидят гидранты, представляют собой как бы продолжение тела полипа. Они также состоят из эктодермы и энтодермы с полостью-каналом внутри, при помощи которого гастральные полости гидрантов сообщаются друг с другом. Снаружи стебли колоний одеты тонкой, прозрачной и твердой органической оболочкой — перидермой, часто образующей перетяжки. У одних полипов перидерма доходит лишь до основания гидранта, тогда как у других она образует вокруг полипа чашечку, или теку, в которую может втягиваться гидрант. На этом признаке основано разделение отряда полипов на подотряды: гидроиды без чашечки, или теки (Athecata), с чашечкой (Thecaphora).
Строение медуз
У многихполипов путем почкования образуются свободноплавающие половые особи — медузы. Тело гидроидных медуз внешне не похоже на гидрантов колонии. Оно может быть сильно выпуклым снаружи и вогнутым внутри, напоминая по форме колокол, или более плоским дисковидным, похожим на зонтик. От центральной вогнутой поверхности зонтика спускается вниз цилиндрический вырост, называемый ротовым хоботком. На его конце расположено ротовое отверстие. По краям зонтика свешиваются щупальца, их может быть 4, 8, 16, редко больше. У основания щупалец или между ними обычно расположены органы чувств. С внутренней стороны края зонтика эктодерма образует кольцевой выступ, так называемый парус или velum.
Медузы плавают, ритмично сжимая и разжимая мускульные волокна края зонтика. При сжатии края зонтика парус прижимается к ротовому хоботку и вода с силой выталкивается из образующейся под зонтиком полости. В этот момент, в силу отдачи, медуза получает реактивный толчок и движется вперед наружной выпуклой поверхностью зонтика. Медузы, будучи мелкими планктонными организмами, передвигаются самостоятельно лишь на незначительные расстояния, но морские течения могут переносить их очень далеко.
Несмотря на кажущееся существенное различие в строении полипов и медуз, последние представляют собой, по существу, измененных полипов с несколько усложненной организацией. Медуза отличается от полипа прежде всего сильным развитием промежуточного вещества — мезоглеи. Если мы представим себе, что у полипа вместо опорной перепонки развивается все утолщающийся слой студенистой мезоглеи, а само тело полипа значительно укорачивается (строго по его продольной оси), причем ротовой конус вытягивается и превращается в хоботок, то нетрудно представить себе превращение полипа в медузу. Разрастание мезоглеи приводит к сближению слоев энтодермы зонтика, кроме центрального участка гастральной полости, расположенного против хоботка. Это сближение слоев энтодермы привело к их срастанию на значительном протяжении. Там, где слои энтодермы не срослись, образовалась система пищеварительных каналов медузы. Рот ведет в желудок, так называют полость внутри ротового хоботка и у некоторых видов уцелевшую центральную часть гастральной полости. От желудка отходят 4, редко больше, радиальных канала, которые соединяются с кольцевым каналом, идущим по краю зонтика. Таким образом, вместо мешковидной гастральной полости полипа у медуз формируется сложная сеть каналов, происходящих из гастральной полости. Она называется гастроваскулярной системой.
Медузы в связи с подвижным образом жизни отличаются от полипов и другими признаками более высокой организации. У медуз дифференцировка эпителиально-мускульных клеток идет значительно дальше. Эпителиальное тело клетки очень невелико по сравнению с ее сильно развитыми мускульными отростками. Такие клетки сконцентрированы вдоль края зонтика и в парусе. Их мускульные отростки вытянуты в длинные волокна, выполняющие функцию кольцевой мускулатуры.
Нервная система у плавающих медуз также организована более высоко, чем у сидячих полипов. Хотя тип ее остается тот же, т. е. диффузной сеткой, или сплетением нервных клеток, но наблюдается дальнейшая концентрация тел нервных клеток по краю зонтика. Их скопления образуют два кольцевых нервных тяжа. Наружное нервное кольцо чувствительное, оно проходит у оснований щупалец, глазков и статоцистов. Внутреннее нервное кольцо двигательное, оно расположено у основания паруса, где концентрируются мускульные волокна.
Кроме осязательных клеток, рассеянных в эктодерме, но более многочисленных по краям зонтика и на щупальцах, у гидромедуз развиты особые органы чувств — глазки и статоцисты.
Органы зрения — глазки — у различных гидромедуз могут быть различной степени сложности. В простейшем случае глазок представляет собой группу удлиненных светочувствительных и пигментных клеток, расположенных в эктодерме. При помощи этого примитивного органа зрения животное только различает свет, но не видит предметов.
Несколько более усложненный орган зрения представляет собой глазная ямка. В этом случае зрительные и пигментные клетки образуют впячивание, причем образовавшееся углубление бывает заполнено студенистым веществом («стекловидное тело»). Если мы представим далее, что глазная ямка отшнуровывается от эктодермы, то образуется глазной пузырь, в центральной части которого дифференцируется светопреломляющее прозрачное тельце, выполняющее функцию хрусталика, а задняя часть пузыря соответствует сетчатке. Это усложнение глаза медуз дает им возможность, по-видимому, лучше ориентироваться в окружающей среде. Глазки всегда располагаются по краю зонтика, у основания щупалец.
Органы равновесия — статоцисты — имеют различное строение, но чаще всего представляют собой замкнутый эктодермический пузырек, лежащий у основания щупальца. Одна из клеток, составляющих стенку этого пузырька, вдается в полость статоциста. В ней помещается известковое тельце статолит. Другие клетки стенки пузырька имеют чувствительные волоски, направленные к клетке со статолитом. К ним подходят отростки нервных клеток от чувствующего нервного кольца. При изменении положения тела медузы статолит в силу закона тяготения надавливает на иную группу волосков чувствительных клеток, чем при нормальном положении тела. Соответствующее раздражение чувствительных клеток передается по их нервным волокнам на внутреннее нервное кольцо, которое контролирует движение паруса. При этом мускульные отростки эпителиально-мышечных клеток паруса на приподнятой стороне зонтика начинают сокращаться более интенсивно до тех пор, пока медуза не примет горизонтального положения. Таким образом, статоцисты осуществляют коррекцию положения медузы в толще воды. Кроме того, по некоторым данным, статоцисты стимулируют ритмичные сокращения краев зонтика (паруса). При удалении статоцистов медуза перестает двигаться, а сокращения краев зонтика становятся крайне редкими.
В заключение можно сказать, что медузы имеют более сложное строение, чем гидранты того же вида Г. полипов. Эти особенности строения объясняются иными условиями жизни свободноплавающих медуз. Именно с этими условиями жизни связана и иная форма тела, и большее развитие мускульных элементов, и особенно развитие нервной системы и органов чувств.
Размножение и чередование покалений
Гидранты у всех морских Г. полипов способны только к бесполому размножению почкованием. При этом путем почкования образуются как гидранты, так и медузоидное поколение. Отличие заключается в том, что гидранты развиваются на стеблях колоний, а медузы отпочковываются на самом гидранте. Однако у некоторых полипов (из подотряда Thecaphora) медузные почки развиваются на видоизмененных гидрантах, лишенных рта и щупалец и заключенных в оболочку — гонотеку, прикрытую крышечкой. Такие особи, образующие медуз, получили название бластостилей.
Медузы являются половой генерацией полипов. Большинство Г. медуз размножается только половым путем. Медузы раздельнополы, и половые железы образуются либо в эктодерме хобота (подотряд Athecata), либо в эктодерме вогнутой части зонтика под радиальными каналами гастроваскулярной системы (подотряд Thecaphora).
Таким образом, у полипов происходит чередование различно размножающихся поколений: бесполого поколения (гидранты) и полового (медузы). Такой тип чередования поколений называют метагенезом.
Биологическое значение смены поколений и образование свободно-плавающего медузоидного поколения заключается в том, что медузки, переносимые течениями на большие расстояния, обеспечивают тем самым расселение вида. Однако у многих Г. полипов чередование поколений осложняется еще тем, что медузы размножаются не только половым путем, но также почкованием, образуя новое поколение медуз. Так, у упомянутой выше медузы Sarsia (Coryne) на хоботке образуются почки, из которых также развиваются медузы.
У многих гидроидных полипов вместо свободноплавающих медуз из медузных почек образуются половые особи, не отрывающиеся от колонии. У различных видов такие особи представляют собой более или менее сильно редуцированных медуз, утративших способность к самостоятельному существованию. Степень упрощения организации остающихся прикрепленными половых особей может быть различна. Если они сохраняют сходство с медузой, но остаются прикрепленными и лишены рта и органов чувств, их называют медузоидами. Если редукция идет далее, их называют гонофорами и споросаками. В простейшем случае споросак представляет собой эктодермический мешочек с энтодермическим стержнем внутри, наполненный созревающими половыми продуктами.
<
Таким образом, половое поколение медуз превращается в своеобразные «половые органы» полиморфной колонии. Как медузы, так и медузоиды, гонофоры и споросаки, образующиеся на одной колонии, формируют половые продукты одного пола: яйцеклетки или сперматозоиды. Зрелые половые клетки освобождаются при разрыве стенок тела медузы или медузоида и выпадают в воду, где и происходит оплодотворение.
Следует отметить, что у некоторых гидроидных полипов в колониях наблюдается не только диморфизм, но и полиморфизм, заключающийся в том, что, кроме типичного гидранта и медузы или медузоида, на ветвях колонии образуются путем почкования еще особые защитные особи — нематофоры, лишенные рта и щупалец, но имеющие большое количество стрекательных клеток.
Развитие
Дробление яйца почти у всех Г. полипов полное и вначале вполне равномерное. У примитивного гидроида Stomateca оно приводит к образованию стадии морулы, представляющей собой беспорядочное скопление эмбриональных клеток — бластомеров. Затем бластомеры зародыша раздвигаются и располагаются по периферии шара; так возникает бластула. На этой стадии зародыш удлиняется, покрывается ресничками и превращается в личинку — целобластулу. У свободноплавающей личинки происходит иммиграция — уход клеток с ее заднего конца внутрь полости бластулы — бластоцеля, или первичной полости тела. Клетки, заполнившие бластоцель, дают начало внутреннему зародышевому листку — энтодерме, а оставшиеся на поверхности — наружному зародышевому листку — эктодерме. Процесс образования двух-слойного зародыша, состоящего из эктодермы и энтодермы, называется гаструляцией. При этом личинка переходит в следующую стадию, называемую паренхимулой, у которой клетки энтодермы сплошь заполняют бластоцель. Позднее клетки энтодермы раздвигаются, и образуется полость первичного кишечника — гастроцеля. При этом паренхимула превращается в планулу — типичную личинку Г. полипов.
Гаструляция у Г. может проходить различными способами. Кроме описанной выше однополюсной (униполярной) иммиграции у Stomateca, различают многополюсную (мультиполярную) иммиграцию, или уход клеток внутрь бластулы с разных ее участков. Кроме того, у некоторых гидроидных (Clava squamata) на стадии морулы наружные бластомеры начинают делиться чаще внутренних и становятся более мелкими. Они отделяются от более крупных внутренних и превращаются в эктодерму, а внутренние — в энтодерму. Этот путь гаструляции называется морулярной деляминацией или расслоением.
Личинка паренхимула или планула, проплавав некоторое время, прикрепляется к субстрату передним расширенным концом, а у некоторых видов боком. На бывшем заднем конце ее прорывается рот и образуются щупальца будущего полипа. Он начинает почковаться, и возникает новая колония.
У гидроида Tubularia яйца содержат значительное количество желтка, что приводит к своеобразной эмбрионизации онтогенеза, или индивидуального развития. Обычные для гидроидных личиночные стадии паренхимулы и планулы проходят у представителей этого рода внутри яйцевых оболочек в гонофоре. Из яйца вылупляется более сложно устроенная личинка с двумя венчиками щупалец, похожая на подвижный полип и называемая актинулой.
Представители отряда морских гидроидов (Leptolida) — типичные обитатели морей и в основном распространены на небольших глубинах. Гидроиды обрастают самые разнообразные подводные предметы — камни, раковины моллюсков, водоросли. Необходимо, однако, иметь в виду, что некоторые представители этого отряда приспособились к жизни в солоноватых и даже пресных водоемах. Очень интересен встречающийся в Каспийском море гидроид Moerisia pallasi. Он обладает двумя типами полипов: плавающими и донными, от которых отпочковываются как полипы, так и медузы.
Геологический словарь
гидра — мифическое чудовище] — класс кишечнополостных (Hydrozoa), объединяет полипов и медуз, обладающих лучевой симметрией тела. Скелет развивается только у некоторых полипов и выделяется эктодермой в виде тонкой роговистой или хитиновой оболочки или пластинки и иногда пропитывается известью. Бесполые поколения (гидроидные полипы) — прикрепляются к субстрату, половые поколения (гидроидные медузы) — свободно плавающие организмы. Первые Г. появи-лись в докембрии. В кембрии уже известны Siphonophora и Stromatoporoidea, Докембрий — современный
Биологический словарь
гидроиды (Hydroidea), подкласс гидроэоев. Объединяет как одиночных полипов и медуз, так и колонии полипов из множества особей (гидрантов). Для мн. Г. характерно чередование поколений полипов и медуз, у нек-рых одно поколение резко доминирует над другим или одно из них отсутствует. 5 отрядов: гидры, хондрофоры, трахилиды, гидрокораллы, лептолиды (по др. системе — 6: гидры, трахилиды, атекаты, текафоры, лимномедузы, актинулиды). Назв. «гидроиды» чаше употребляют для обозначения только гидроидных полипов; относящихся к этим полипам медуз наз. гидроидными медузами (или гидромедузами).
Поделиться с друзьями