
Губки тип классы
Губки — наиболее низкоорганизованные асимметричные или радиально-симметричные многоклеточные животные (табл. I, рис. 57). Тип губок включает около 5000 видов, подавляющее большинство которых обитает в морях и океанах, пресноводных видов немного (сем. Spongillidae).
Систематическое положение - Лучистые или Радиально-смметричные. Латинское Radialia
Тип Губки Латинское название Porifera или Spongia
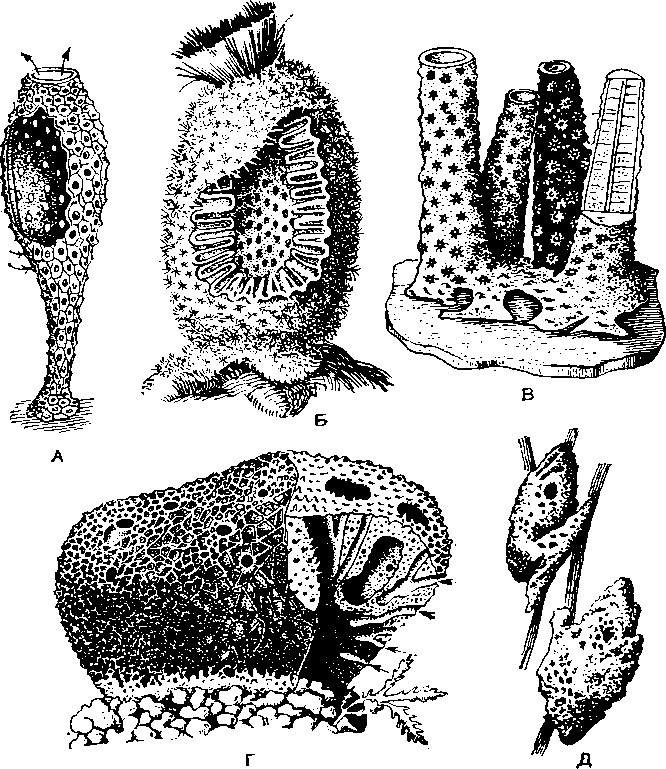
А — Leucosolenia coriacea; Б — Sycon raplianus: В — колония Verongia aerophoba; Г — туалетная губ. (Spongia officinalis): Д — пресноводная бадяга Spongilla.
Общая характеристика губок
Все губ. во взрослом состоянии неподвижные, прикрепленные животные, не имеющие ни нервных клеток, ни органов чувств, ни мускулатуры, ни ротового отверстия. Постоянный ток воды со взвешенными пищевыми частицами проходит через все тело животного. Вода попадает через многочисленные поры (Porifera означает — несущие поры), расположенные на наружной поверхности тела губок, в систему поровых каналов и жгутиковых камер, или полостей, и выводится наружу через устье, или оскулум.
Губки находятся на клеточном уровне организации. Это означает, что их тело представляет собой скопление рыхло расположенных, в разной степени дифференцированных клеток, выполняющих определенные функции: покровную, опорную, вододвигательную, питательную и др. Однако эти клетки еще не образуют настоящих тканей и органов.
У губок различают следующие основные типы клеток:
- Пинакоциты — плоские многоугольные, способные сокращаться клетки, которые выстилают наружную поверхность и поровые каналы Г..
- Воротничков ые клетки, или хоаноциты, цилиндрической или шаровидной формы. Они образуют жгутиковые камеры, а у наиболее примитивных одиночных известковых губок выстилают внутреннюю (гастральную) полость. На свободном конце клетки, обращенном внутрь жгутиковой камеры, расположен жгутик, окруженный цитоплазматическим воротничком (рис. 2). С помощью электронного микроскопа выяснено, что воротничок хоаноцита образован венчиком из 30—40 тесно прилежащих друг к другу тончайших плазматических выростов — щупалец. Жгутик хоаноцита отходит от базального тельца, к которому присоединено чешуевидное парабазальное тельце (рис. 2, В). Такое же ультратонкое строение имеют и воротничковые жгутиковые (Craspedomonadina), что указывает на их филогенетическую близость. Полагают, что Г. и воротничковые жгутиковые имели общего предка. Хоаноциты выполняют вододвигательную и питательную функцию. Биение их жгутиков создает постоянный направленный ток воды в теле Г., а пищевые частицы, приносимые водой, захватываются и перевариваются хоаноцитами. Весь процесс пищеварения у губок проходит внутриклеточно. Непереваренные остатки пищи из пищеварительных вакуолей выбрасываются с током воды через оскулум.
- Амебоциты — крупные блуждающие клетки, они осуществляют фагоцитоз и участвуют в переваривании пищи, получая излишек пищевых частиц от хоапоцитов. Амебоциты постоянно передвигаются в студенистой массе основного вещества Г. — мезоглее, которая заполняет все пространство между поверхностью Г. и системой водоносных каналов. Мезоглея связывает все типы клеток механически и химически, или гуморальн
- Колленциты — звездчатые клетки, соприкасающиеся своими отростками с соседними клетками, они выполняют опорную функцию.
- Склеробласты и сионгиобласты — крупные клетки, формирующие элементы скелета Г. — спикулы и спонгинотше волокна.
- Археоциты — не дифференцированные клетки, способные легко превращаться в клетки любого типа, служат клеточным резервом.
- Половые клетки, образующиеся из археоцитов, обеспечивают половое размножение.
Все типы клеток губок способны при некоторых условиях переходить в амебоидное состояние и превращаться в клетки иного типа. Даже рано обособившиеся хоаноциты, по последним данным, способны терять воротнички и жгутики, уходить в глубь мезоглеи и превращаться там в амебоциты. Губ. можно разрезать на маленькие кусочки и протереть через мелкоячеистое сито (мельничный газ), при этом клетки будут отделены друг от друга. Если протертую массу поместить на дно сосуда, то клетки будут сползаться и образовывать группы, или агрегаты, клеток, которые через несколько дней превратятся в маленьких губок. Такая высокая р е-ге нерационная способность губок объясняется относительно слабой дифференцировкой их клеток и отсутствием настоящих тканей.
Г., как и все многоклеточные, размножаются половым путем. Формирование половых клеток — яйцеклеток и сперматозоидов — происходит обычным для всех многоклеточных способом. Кроме полового размножения, губ. присуще бесполое размножение — путем наружного почкования или образования покоящихся внутренних почек — геммул.
Внешний вид и внутреннее строение
Губ. совсем не похожи на животных, они скорее напоминают растения. Недаром их до начала XIX в. относили к сборной группе зоофитов, будто бы промежуточной между растениями и животными. Форма губок крайне разнообразна. Многие из них образуют наросты и корки на субстрате, караваи, глыбы. Немало губок имеет чашевидную, кустистую, трубчатую, грибовидную (рис. 1, А, Б, В; табл. I) форму. Неопределенность формы тела многих губок зависит от того, что они образуют колонии, состоящие из многих особей. При разрастании колонии обычно теряет индивидуальность составляющих ее особей (рис. 1, Г, Д). Отдельные особи ясно различимы только у немногих колониальных губок (рис. 1, В; табл. I). Одиночных губок сравнительно немного. Обычно они имеют более правильную форму — вазы, кубка или бочонка. У одиночных губок можно выявить радиальную многолучевую симметрию.
На ощупь тело большинства губок жесткое, что объясняется присутствием твердого скелета — известкового, кремневого или кремнерогового.
Размеры губок колеблются от нескольких миллиметров до 1—1,5 м в длину и до 2 м в диаметре. Многие морские виды ярко окрашены, чаще в желтые, оранжевые, красные, пурпурно-фиолетовые и коричневые тона. Окраску обусловливают разнообразные пигментные клетки, присутствующие в мезоглее. Некоторые гГ. лишены пигмента и выглядят белыми или сероватыми. Пресноводные Г. бадяги окрашены в коричневатый или зеленый цвет. Последний зависит от присутствия симбиотических одноклеточных зеленых водорослей — зоохлорелл.
Большинство губок имеет резкий, неприятный запах, который зависит от выделения ядовитых веществ. Само тело губок также обладает токсическими свойствами, имеющими, по-видимому, защитное значение и обусловливающими наряду с твердым скелетом несъедобность губок. Предполагают, что запах и токсичность губок обусловлены накоплением значительного количества продуктов метаболизма в отдельных клетках и в мезоглее губ.
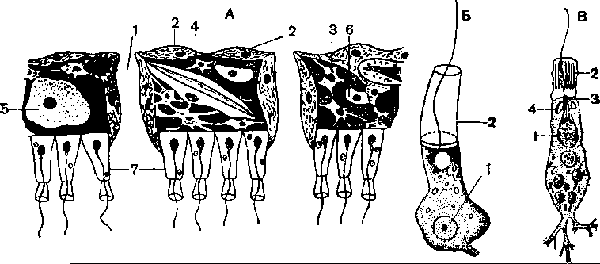
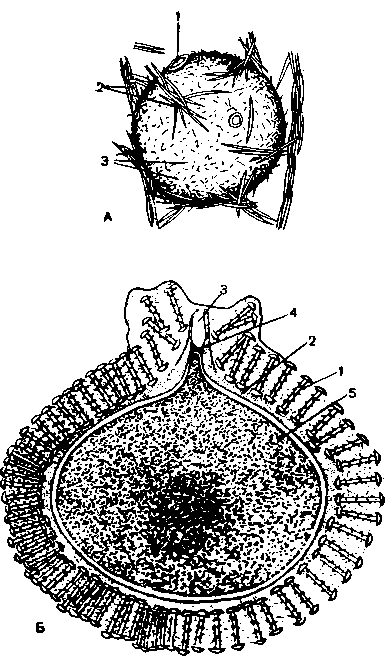
А разрез через часть стенки тела Г. асконоидного типа; 1 — поровый капал; 2 — пина-коциты; 3 — звездчатые клетки мезоглеи; 4 — склеробласт с образующейся внутри клетки спикулой; 5 — яйцевая клетка; 6 — археоцпт; 7 — хоаноциты; Б — хоаноцит; В — ультратонкое строение хоаноцита известковой Г. r.Sycon raplianus); 1—ядро; 2 — плазматический воротничок; 3 — баззльксе тельце; 4 — парабазальное тельце.
Внутреннее строение губок легче всего изучать на одиночных формах. При этом различают три типа их организации: а с коноидный, или аскон, сиконоидный, или сикон, и лейконоидный, или лейкон.
Наиболее просто устроены очень маленькие одиночные (по крайней мере молодые) известковые Г. Leucosolenia coriacea (рис. 1, А; рис. 3, А). Их радиально симметричное тело похоже на вазу. Тонкая стенка тела (около 2 мм) ограничивает центральную гастральную, или атриальную, полость губ.. Последняя одета изнутри почти сплошным слоем хоаноцитов, образующих примитивный рыхлый жгутиковый эпителий, который свидетельствует о зачаточном тканеобразовании у губок. Наружпая дермальная поверхность тела покрыта преимущественно пинакоцитами (рис. 2, Л), перемешанными с клетками других типов. Весьма многочисленны трубкообразные пороциты, соединяющие наружную поверхность гГ. с гастральной полостью. Их внутриклеточный. просвет образует поры, по которым вода поступает в гастральную полость. Между дермальной поверхностью и слоем хоаноцитов в студенистом веществе мезоглеи расположены многочисленные звездчатые колленциты, склеробласты со спикулами и образованные ими трехосные спикулы, различной формы амебоидные клетки — амебоциты и археоциты, веретеновидные сократимые клетки
— миоциты и др.
<
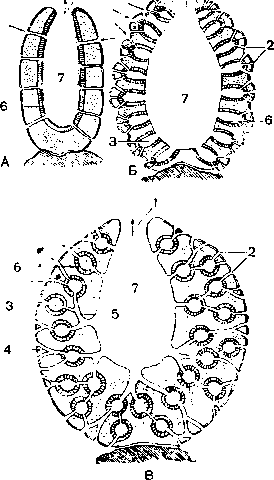
А — асконоидный; Б — сиконоидный; В — лейконоидный; 1 — оскулум; 2 — поры; 3 — жгутиковые камеры; 4 — приносящие каналы; 5 — выносящие каналы; 6 — хоаноциты; 7 — гастральная полость. Стрелки показывают направления . токов воды.
Непрерывное биение жгутиков хоаноцитов прогоняет воду из гастральной полости через выводное отверстие — оскулум — наружу, а через поры в тело Г. поступают все новые порции воды.
Этот тип организации называется аскон, он встречается у небольшого числа видов известковых губок или у молодых губок, строение которых во взрослом состоянии усложняется. Большинство губок устроено сложнее.
Усложнение организации губок (тип сикон) наблюдается у небольших известковых губок из рода Sycon и др. (рис. 1, Б; 39, Б). Стенка тела этих губок утолщается, в ней образуются многочисленные цилиндрические впячивания атриальной полости. Хоаноциты располагаются только в этих впячиваниях, которые превращаются в жгутиковые каналы. Атриальную полость сиконоидной Г. выстилают только пина-коциты. Вода поступает через межклеточные поры в норовые каналы, выстланные пинакоцитами, затем в жгутиковые каналы, а оттуда в атриальную полость и выводится через оскулум наружу.
Большинству крупных одиночных и колониальных губок свойственно дальнейшее усложнение организации. При этом жгутиковые каналы разбиваются на маленькие шарообразные жгутиковые камеры, которые лежат в несколько слоев внутри еще более утолщенной стенки тела Г.. Все жгутиковые камеры связаны с внешней средой сложной системой приводящих каналов, а с атриальной полостью — системой отводящих каналов. Этот наиболее сложный тип строения называют лейконоидным (рис. 59, В). Прогрессивное увеличение числа маленьких жгутиковых камер приводит к значительному повышению их фильтрационной способности. Так, у небольшой Г. Leucandra aspera, высотой 7 см и диаметром 1 см, подсчитано 2 250 000 жгутиковых камер с общей поверхностью 52,5 см2, 81 000 приносящих и 5200 отводящих каналов. За сутки такая губ. фильтрует 22,5 л воды.
У многих губок вторично образуются защитные дермальные мембраны, или корковый слой, покрывающий снаружи тело губ.. Атриальная полость может сильно сокращаться, мало отличаясь от отводящих каналов, особенно у колониальных форм. У последних обычно невозможно различать отдельные особи, входящие в состав колонии. Об их количестве можно судить лишь по числу оскулумов Г..
Питание губок осуществляется хоаноцитами и амебоцитами. Хоа-ноциты захватывают мелкие пищевые частицы (детрит, бактерии, одноклеточные водоросли и т. п.) короткими псевдоподиями, образующимися несколько ниже воротничка. При этом формируются типичные пищеварительные вакуоли, совершающие круговые движения в цитоплазме хоа-ноцита. Непереваренные остатки пищи выбрасываются из сократившихся в размерах вакуолей в зоне воротничка. Крупные хоаноциты известковых губок захватывают пищевые частицы, а затем передают их прилегающим к ним амебоцитам, которые их переваривают и разносят питательный материал по телу. Более крупные частицы фагоцитируются непосредственно амебоцитами, подходящими к стенкам каналов.
Дыхание и выделение у губок осуществляются диффузно. Кислород, растворенный в воде, которая проходит через все тело губ., используется ею для дыхания (газообмена). Жидкие продукты метаболизма и экскременты из пищеварительных вакуолей хоаноцитов и амебоцитов поступают через систему каналов в атриальную полость, а оттуда с током воды выбрасываются через оскулум наружу.
До настоящего времени не выяснено, каким путем осуществляется координация между всеми клеточными элементами губок. Французские ученые (Тюзе, Паван, Грассе и др.) описали биполярные и мультиполяр-ные паукообразные и пузыревидные клетки, образующие связи с клетками других типов. Они считают их за нервные клетки, образующие диффузную нервную систему. Однако у губок крайне низка способность реагировать даже на грубые внешние раздражения (химические, термические и др.). Губ. можно проткнуть раскаленным металлическим стержнем, однако реакция на это раздражение будет распространяться по телу очень медленно и на незначительное расстояние. Все движения взрослой губ. сводятся к медленному закрыванию пор и оскулума. Все это дает основание большинству ученых отрицать наличие у губок как нервной, так и мышечной системы.
Скелет
Подавляющее большинство губок имеет скелет. Особенности в строении и составе скелетных образований являются важнейшим систематическим признаком. Скелет у различных групп губок может быть известковым, кремневым, кремнероговым и роговым, или спонгиновым.
Известковый скелет свойствен сравнительно небольшой группе исключительно морских, чаще одиночных губок (Leucosolenia, Sycon и др.). Он образован различными по форме иглами, или спикулами, состоящими из углекислого кальция. Различают одно-, трех- и четырехлучевые иглы (рис. 4, А). У сикона и других губок (рис. 57) они окружают жгутиковые камеры и оскулум, причем концы их могут торчать наружу.
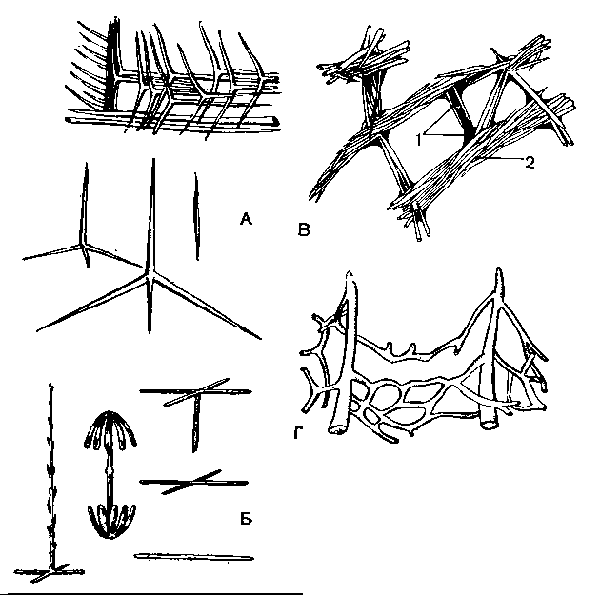
А — спикулы известковых губок; Б — кремневые спикулы стеклянных губок; В — скелет бадяги; 1 — спонгиновые волокна; 2— кремневые спикулы; Г — спонгиновый скелет туалетной губки.
Спикулы образуются особыми клетками мезоглеи — склеробла-стами. Отложение углекислого кальция и начало образования спикулы происходит внутри склеробласта. Когда спикула достигает размеров, превышающих размеры склеробласта, последний отмирает, а игла остается в мезоглее.
Формирование трех- и четырехлучевых спикул происходит при совместной деятельности нескольких склеробластов, образующих однолучевые спикулы, которые одновременно спаиваются своими концами под определенным углом.
Кремневый скелет образован спикулами, состоящими из кремнезема, который отлагается в склеробласте вокруг осевой нити. Последняя состоит из органического вещества.
Кремневые спикулы бывают двух родов: более крупные — макросклериты, составляющие основу скелета, и мелкие — микросклериты. Кремневые спикулы весьма разнообразны по форме (рис. 60, Б). Макросклериты могут быть одно-, трех- и четырехосными, достигая у некоторых губок больших размеров. Так, у стеклянной Г. Hyalonema (рис. 63, Б) «корневые» нити имеют длину в несколько десятков сантиметров, а у губ. Monorhaphis так называемая осевая спикула достигает длины 3 м
Кремнероговой скелет имеется у бадяг и некоторых других губок (рис. 4, В). Он состоит из кремневых спикул и волокон, образованных особым органическим веществом — спонгином. Спонгин — сложное органическое вещество, по составу близкое к шелку, содержащее иод (до 14%). Волокна спонгина образуются не внутри отдельных клеток, а межклеточно. Мелкие клетки спонгиобласты как футляром одевают растущее волокно.
Роговой, или спонгиновый, скелет, свойственный небольшому числу губок (туалетная губ.), вследствие редукции кремневых спикул состоит только из спонгина (рис. 60, Г).
Некоторые очень мелкие губ. полностью лишены скелетных образований. По-видимому, утеря минерального скелета роговыми и бесскелет-ными губками произошла вторично.
Губкам свойственно как бесполое, так и половое раз-Размпожение множение. Бесполое размножение губок проис-и развитие ходит двумя способами: наружным и внутренним почкованием.
Наружное почкование заключается в образовании небольших вздутий — почек, затем разрастающихся и формирующих собственный оскулум. У большинства губок дочерние особи не отделяются от материнской, и образуется колония. У одиночных губок дочерние особи отделяются от материнской и переходят к самостоятельному существованию.
Внутреннее почкование наблюдается у пресноводных бадяг и некоторых морских губок. У бадяг осенью археоциты собираются в группы и вокруг них образуется двойная хитиноидная оболочка. Наружный слой этой оболочки имеет воздушные камеры. Склеробласты, окружающие внутреннюю почку, или гем му л у (рис. 61), образуют особые склериты, так называемые амфидиски, обычно состоящие из колонок с звездообразными расширениями на концах. Амфидиски располагаются между двумя слоями оболочки.
Осенью колонии бадяг погибают, а внутри их сохраняются жизнеспособные геммулы. Они хорошо переносят промерзание и высыхание водоема. Геммулы могут оставаться на месте старой колонии или, при разрушении ее, переноситься течением, ветром или водоплавающими птицами. Таким образом, геммулы обеспечивают как переживание губок в течение неблагоприятного времени года, так и их расселение.
Весной из геммул выходят археоциты и образуют новую губ., причем содержимое нескольких геммул часто сливается вместе и развивается в одну колонию.
А — геммула озерной бадяги (Spongilla lacust-ris); 1 — поровое отверстие; 2 — мегасклеры; 3 — микросклеры; Б — разрез через геммулу бадяги Ephydatia blembingia; 1 — амфидиски; 2 — внутренняя кутикулярная оболочка; 3 — поровое отверстие; 4 — замыкающая его перегородка; 5 — внутренняя масса клеток.
Половоеразмножение свойственно всем губ. Значительная часть губок — гермафродиты, но многие, в том числе пресноводные бадяги, раздельнополы. У раздельнополых губок одни колонии образуют только яйцевые клетки, а другие — только сперматозоиды. Половые клетки образуются в мезоглее из археоцитов. Женские половые клетки — яйца, или ооциты, остаются в мезоглее, где и происходит их оплодотворение. Мужские половые клетки — сперматозоиды — выходят в воду и проникают с током воды в соседние колонии.
Оплодотворение у всех губок проходит очень своеобразно. Сперматозоид проникает через воротничок внутрь хоаноцита, где теряет хвост и инцистируется. Такой хоаноцит со сперматозоидом внутри переходит в амебоидное состояние и превращается в клетку носительницу. Последняя мигрирует в мезоглею, где поглощается ооцитом. При этом сливаются гаплоидные ядра ооцита и сперматозоида, т. е. происходит процесс оплодотворения.
Дробление оплодотворенного яйца у губок происходит обычно внутри материнского организма, где образуются личинки. Развитие у губок протекает очень разнообразно и подчас весьма сложно.
У некоторых известковых губок (Sycon raphanus) дробление яйца проходит в мезоглее под слоем хоаноцитов (рис. 6, А). В результате его образуется бластула, у которой восемь крупных клеток тесно прилегают к хоаноцитам, а мелкие, более многочисленные жгутиковые клетки составляют ее вторую часть, расположенную свободно в мезоглее. Жгутики мелких клеток направлены внутрь полости бластулы. Затем в центре бластулы раздвигается группа крупных клеток и образуется отверстие, через которое становятся видны усиленно размножающиеся жгутиковые клетки. Внутренняя часть личинки со жгутиковыми клетками выгибается в виде купола по направлению к слою хоаноцитов, а крупные клетки отходят от хоаноцитов, располагаются по краям отверстия, а затем смыкаются на полюсе, противоположном их первоначальному положению, где ранее были мелкие клетки. Этот процесс называется экскурвацией или вы-ворачиванием поверхностей зародыша. Такого изменения, или инверсии, поверхностей бластулы у других многоклеточных не наблюдается. У простейших аналогичный процесс известен только при формировании колоний вольвокса. Крупные клетки впячиваются при этом внутрь образовавшейся амфибластулы (рис. 62, А). Затем они вновь выпячиваются, и шаровидная личинка покидает материнский организм. Амфи-бластула свободно плавает за счет биения жгутиков, расположенных на ее переднем полюсе, а задний полюс образован крупными зернистыми клетками (рис. 6, А).
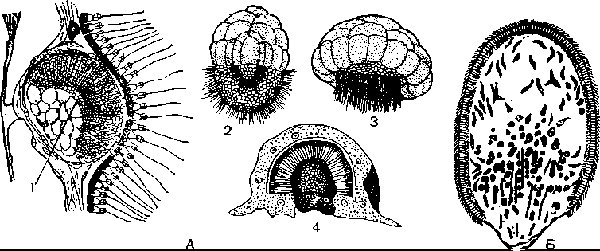
А— развитие губки Sycon raphanus; l — амфнбластула в теле материнской губ.и (видно впя-чивание крупных клеток); 2 — плавающая амфнбластула; 3 — впячпвание мерцательных клеток; 4 — прикрепившаяся личинка; Б — личинка кремнероговой губ. МухШа.
После короткого периода подвижного существования амфнбластула прикрепляется передним полюсом к поверхности дна. При этом мелкие жгутиковые клетки впячиваются внутрь полости амфибластулы, а крупные зернистые образуют наружный слой, т. е. происходит процесс гаст-руляции. Бластопор, которым личинка прикрепилась к субстрату, закрывается, жгутиковые клетки теряют жгутики (де дифференцируются) и усиленно размножаются. Позднее из них образуются хоаноциты жгутиковых каналов, а наружные крупные зернистые клетки превращаются во все остальные клетки тела Г. (рис. 6, А).
У некоторых обыкновенных губок, например у Oscarella lobularis и других, в материнском организме развивается однослойная бластула — целобластула, равномерно покрытая жгутиками. Она покидает материнскую губ. и свободно плавает. Ее задний полюс образован более длинными красноватыми клетками, а клетки переднего полюса бесцветные. Затем она садится на дно передним полюсом, бесцветные клетки которого впячиваются внутрь, т. е. происходит типичная гаструляция путем инвагинации. Наружные красные клетки теряют жгутики, часть из них распадается. Бластопор закрывается, и происходит сложный процесс формирования жгутиковых камер за счет жгутиковых клеток переднего полюса, все остальные части тела Г. (система каналов, выстилка наружной поверхности и атриальной полости, скелет и т. п.) образуются путем размножения и дифференцировки клеток заднего полюса целобластулы.
У части обыкновенных губок (Monaxonida, Keratosa) губку-мать покидает более сложно организованная личинка — паренхимула (рис. 62, Б). Снаружи она почти целиком покрыта жгутиковыми клетками (у некоторых видов на заднем полюсе нет жгутиков), а внутри ее находятся рыхло расположенные клетки, которые у отдельных видов способны дифференцироваться на амебоциты и склеробласты. Еще до вылупления личинки из материнской губ. в ней начинается формирование спикул.
После короткого периода свободного плавания паренхимула прикрепляется передним концом к дну и сильно уплощается. Все жгутиковые клетки ее мигрируют внутрь тела (поодиночке или группами), где их жгутики исчезают. Позднее эти клетки превращаются в хоаноциты и образуют жгутиковые камеры. Одновременно внутренние клетки обрастают всю наружную поверхность Г.. Часть внутренних клеток мезоглеи выстилает приводящие и отводящие каналы, атриальную полость, прорывающийся оскулум. Этот способ развития губ. соответствует поздней гаст-руляции путем мультиполярной иммиграции и эпиболии прочих многоклеточных.
У пресноводных губок бадяг (Spongilla lacustris) наблюдается еще большая эмбрионизация развития. В материнском организме губ. в паренхимуле происходит формирование не только амебоцитов и склеробла-стов, но и жгутиковых камер. Такую паренхимулу правильнее считать молодой губ., еще не имеющей системы каналов. Она появляется из оскулума матери, покрытая жгутиковыми клетками. После ее фиксации на грунте водоема жгутиковые клетки мигрируют внутрь мезоглеи, где их фагоцитируют амебоциты.
Таким образом, в данном случае жгутиковые клетки выступают как временный личиночный орган движения. Дальнейшее развитие такой губ. сводится к построению системы каналов, выстилке дермальной поверхности, атриальной полости и т. п.
Некоторые ученые рассматривают первое впячивание крупных клеток амфибластулы известковых губок, проходящее в материнской губке, как истипную гаструляцию. Тогда впячивание мелких жгутиковых клеток ее заднего полюса после оседания амфибластулы на дно будет вторичным, а не настоящей гаструляцией. Соответственно эти ученые считают губок животными с вывернутыми слоями. При развитии обыкновенных губок поверхностные жгутиковые клетки паренхимулы мигрируют внутрь и превращаются в пищеварительные клетки — хоаноциты. Это тоже как бы подтверждает вывертывание зародышевых слоев тела, в котором видели коренное отличие губок от всех остальных многоклеточных животных. Более подробное изучение эмбрионального развития губок показало, что у некоторых форм, описанных выше, никакого выворачивания слоев не происходит. Более того, у пресноводных губок поверхностные и жгутиковые клетки играют роль провизорного личиночного оргапа передвижения и в построении тела взрослой Г. участия не принимают.
Поверхностные жгутиковые клетки личинок губок можно рассматривать как кин облает — двигательный слой. При переходе к сидячему образу жизни он у взрослой губ. уходит внутрь и выполняет вододвигательную функцию. Фагоцитобласт — внутренний слой — дает все типы клеток губок, как типичных фагоцитов, так и выстилающих пограничные поверхности (наружную дермальную поверхность и систему каналов). Только в этом смысле можно говорить об извращении первичных слоев у губок. Никакой экто- и энтодермы, т. е. настоящих покровных и пищеварительных тканей, у губок еще нет, как нет у них и настоящих зародышевых листков. О крайне низком уровне клеточной интеграции свидетельствует и разнообразие типов их индивидуального развития.

Классификация
Основным признаком, на котором основывается классификация губок, служит строение их скелета. Тип губок разделяется на три класса: 1. Известковые губки (Calcarea); 2. Стеклянные губки (Hyalospongia); 3. Обыкновенные губки (Demospongia).
По современной классификации:
Тип/Отдел Porifera • 9,092 из предполагаемых 8,659 видов (100%) и ископаемые
Класс Calcarea - Известковые губ. • 775 современных вида; и ископаемые
Класс Demospongiae - Обыкновенные губ. • 7,532 современных вида; и ископаемые
Класс Hexactinellida Шестилучевы́е гу́б., или стекля́нные гу́б. • 664 современных вида и ископаемые
Класс Homoscleromorpha • 120 современных вида;
Класс Not assigned • 1 современный вид
Класс Calcarea - Известковые губки
Известковые губ. — исключительно морские мелкие, часто одиночные формы. Их скелет образован известковыми спикулами. Обычно это обитатели мелководий: Leucosolenia, Sycon, Sycetta, Leuconia и др« (рис. 1, А, Б).
Класс Hexactinellida Шестилучевы́е гу́бки, или стекля́нные гу́бки
К этому классу относятся также только морские, но глубоководные губ. с кремневым скелетом, образованным из спаянных вместе трехосных спиьул (макросклеритов). К ним принадлежат Eupiectella (рис.7, А), Hyalonema (рис. 7, Б) с пучком длинных хвостовых кремневых игл, которыми они удерживаются в илистом грунте, Monorhaphis и др.
Класс Demospongiae - Обыкновенные губки
Наиболее сложно организованные (лейконоидные) губки с кремневым скелетом из четырехосных или одноосных макросклеритов, часто укрепленных спонгиновыми волокнами. Обыкновенные Г. наиболее многочисленны и разнообразны. Этот класс включает несколько отрядов.
Скелет этих морских губок образован исключительно четырехосными спикулами. В Баренцевом море в большом количестве встречается Geodia (табл. I), образующая крупные шаровидные колонии, до 50 см в диаметре. Некоторые представители этого отряда — Oscarella, Halisarca — лишены спикул; их называют мясистыми губками.
ОТРЯД 2. ОДНООСНЫЕ ГУБКИ (MONAXONIDA)
Скелет этих губок образован одноосными кремневыми спикулами, связанными сетью спонгиновых волокон в прочную массу. У некоторых видов спикул становится мало, а спонгиновых волокон больше.
Одноосные крайне разнообразны, к ним относятся как морские, так и пресноводные виды. Среди морских губок известен кубок Нептуна (Poterion neptuni), который обитает в Тихом океане и достигает 1,5 м. Интересны также сверлящие губ. (Cliona), поселяющиеся в известняках, на раковинах моллюсков и на коралловых полипах.
Большая группа широкораспространенных пресноводных губок бадяг (сем. Spongillidae) образует корки или массивные бесформенные наросты (рис. 57, Д) на дне водоемов (на камнях, стеблях водных растений и т. п.), часто окрашенные в зеленый или бурый цвет. Байкальские Г. (табл. I) на небольших глубинах образуют кустистые заросли до 1 м высотой.
ОТРЯД 3. РОГОВЫЕ ГУБКИ (KERATOSA)
Обыкновенные губки, скелет которых не содержит кремневых спикул, а образован только сетью спонгиновых волокон. Обычно массивные крупные формы: конская губка (Hyppospongia communis), достигающая 90 см в диаметре, веронгия (Verongia, рис. 57, В) и др. Наибольшее значение имеет туалетная губка (Spongia officinalis), обычная в Средиземном море, которая издавна служит предметом промысла (рис. 57, Г).
Класс Homoscleromorpha
Отряд морских губок, выделяемый в монотипический класс Homoscleromorpha, ранее рассматривавшийся в составе класса обыкновенных губок. Цитологическая организация отличается от всех остальных губок. ... По современным филогенетическим данным, Homoscleromorpha — чётко ограниченная монофилетическая группа губок, независимая от обыкновенных губок. По данным сайта World Porifera Database, на июль 2016 года в отряд включают следующие таксоны до рода включительно: Семейство Oscarellidae Lendenfeld, 1887.
ФИЛОГЕНИЯ ГУБОК
Губ.. — очень своеобразная группа многоклеточных животных. По уровню организации они занимают промежуточное положение между простейшими и кишечнополостными. Общность ультратонкого строения во-ротничковых клеток губок и воротничковых жгутиконосцев Craspedomo-iiadina говорит о том, что они произошли от одного предка.
Будучи типичными многоклеточными животными, Г. обладают некоторыми чертами простейших:
1. Хоаноциты расположены рыхло, они могут уходить в мезоглею и переходить в амебоидную форму.
2. Многие типы клеток губок имеют амебоидную природу. Так, женские половые клетки — ооциты — двигаются при помощи псевдоподий, вылезают в атриальную полость в поисках питательных клеток и уползают обратно.
3. При оплодотворении ооцита в нем образуется углубление — клеточный рот; он захватывает инцистированный сперматоцит, как амеба захватывает добычу.
4. Процесс инверсии поверхностей, имеющий место при развитии ам-фибластулы известковых губок, известен только у простейших (Volvoci-dae). Последнее вовсе не говорит о родстве губок и вольвоксовых. По-видимому, губ. произошли от колоний примитивных воротничковых жгутиковых, подобных вольвоксу.
[sch:11]
Губ. ничем принципиально не отличаются от остальных многоклеточных животных. Их хоаноциты, или пищеварительные клетки, располагаются в один слой, как и у остальных Metazoa. Воротничковые клетки имеются и у других многоклеточных: у некоторых иглокожих (морские ежи, голотурии) и кишечнодышащих. Образование половых клеток (сперматогенез и оогенез) и строение сперматозоидов не отличаются от таковых даже высокоорганизованных многоклеточных. У некоторых обыкновенных губок (Demospongia) ^совсем не происходит выворачивания первичных (зародышевых) слоей При развитии известковых губок проявляется четырехлучевая симметрия (четыре крупные клетки образуют крест), весьма характерная для кишечнополостных, личинок полихет, га-стропод и сипункулид. О родстве губок с остальными многоклеточными свидетельствует также сходство личинок губок с личинками кишечнополостных и иглокожих.
Все приведенные сведения подтверждают теорию общего происхождения губок и остальных многоклеточных.
Отсутствие нервной и мышечной систем у губок говорит об их примитивности. Губок можно поставить у самого начала общего ствола Metazoa.
Поделиться с друзьями
