
Экология рептилий - Пресмыкающиеся
Условия существования и общее распространение
Пресмыкающиеся живут в значительно более разнообразных условиях, чем амфибии. Это прямым образом связано с более высоким уровнем организации пресмыкающихся, обусловившим возможность широкого их распространения в различных жизненных средах, и в первую очередь в наземной среде. Одной из наиболее важных в этом смысле особенностей является их способность переносить сухость наземной среды как во взрослом состоянии, так (что особенно важно) и в период эмбрионального развития. У взрослых животных это связано с уже описанным ороговением эпидермиса. Эмбриональные приспособления будут описаны ниже, в разделе о размножении.
Возможность жить и размножаться на суше позволила рептилиям приспособиться к обитанию не только в условиях теплого, влажного климата, но и в обстановке сухих, жарких пустынь. В итоге общее географическое распространение пресмыкающихся значительно более широко, чем амфибий. Они заселяют все климатические зоны земного шара, за исключением арктической и антарктической. Вместе с тем надо учесть, что в связи с непостоянной температурой тела рептилии успешно существуют только при относительно высокой температуре среды. Так, например, змеи уже при +10° С становятся малоактивными, при +6, +8° С они перестают двигаться, а при температуре +2, +3° С впадают в оцепенение. Охлаждение тела до —4, —6° С вызывает смерть.
Соответственно сказанному рептилии наиболее многочисленны в тропическом поясе, а по направлению к полюсам обилие видов и особей становится все меньшим.
Так, на островах Индонезии живет 150—200 видов, а в Среднем Китае всего лишь около 30.
У нас в Средней Азии обитает около 50 видов, на Кавказе — 68, в средней полосе Западной Европы — 12, наконец, до Северного полярного круга в западной (наиболее теплой) части Европы доходят только 2 вида (гадюка и живородящая ящерица). Также заметно сокращается обилие рептилий в горных странах по мере поднятия вверх над уровнем моря. Верхний предел распространения рептилий в Центральной Азии лежит на высоте около 5000 м над уровнем моря.
В Кордильерских горах Южной Америки один из видов ящериц (Liolaemus multiformes) распространен до высоты 4900 м над уровнем моря.
Вместе с тем чрезмерно высокая температура для рептилий, естественно, губительна. Привязанные за лапку такырные круглоголовки в пустыне Бетпак-Дала при температуре почвы 46—48° С чувствовали себя весьма стесненно. Дыхание их резко учащалось; они широко раскрывали рот и старались влезть на какое-либо возвышение, где температура воздуха была ниже и большая подвижность воздуха способствовала охлаждению тела. Ящерица (Eremias), находящаяся в таком же состоянии на песке, температура которого на поверхности была равна 55° С, первоначально реагировала подобным же образом, но через 1,5—4 минуты организм не выдерживал перегревания, и животное погибало (Кашкаров). Спасаясь от перенагревания, ящерицы в наиболее жаркую часть дня залегают в норы (агамы) или залезают на ветки кустарников. Д. Н. Кашкаров установил, что в песчаных пустынях Средней Азии летом в середине дня температура воздуха на ветке на высоте 2 м над почвой была на 10° С ниже температуры почвы в тени и на 28° С ниже температуры почвы на солнце.
Высокая температура оказывает и косвенное отрицательное влияние на жизнедеятельность рептилий. Выгорание растительности вызывает летнюю спячку степных черепах. В тропиках высокая температура и пересыхание водоемов вызывают спячку у крокодилов, некоторых черепах и змей.
Наличие в воде или почве солей не мешает существованию рептилий. Многие виды ящериц и змей живут на сильно засоленных почвах, где обитание амфибий совершенно невозможно. Змеи, черепахи (в том числе и лишенные рогового панциря) долгое время без вреда для организма проводят в соленой воде морей и озер.
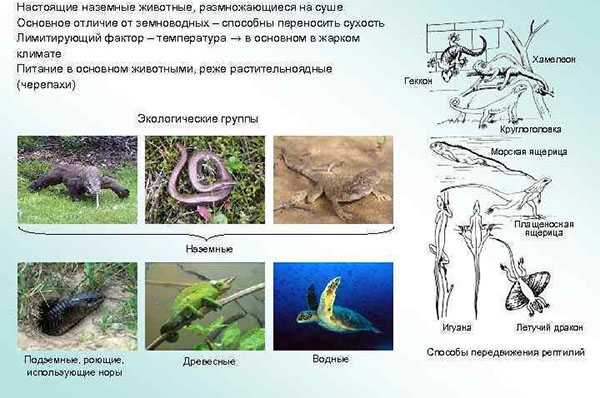
Общее экологическое распространение пресмыкающихся исключительно широко. Среди них есть виды наземные, полуводные, водные, подземные, живущие на деревьях. Современные рептилии отсутствуют только в воздухе. Такое разнообразие жизненных форм понятно, если учесть, что живущие ныне рептилии представляют остатки многих, далеко разошедшихся ветвей, представители которых издавна были приспособлены к обитанию в весьма разнообразных условиях.
Наибольшее число видов принадлежит к наземным животным. Однако и в этом случае видно, что рептилии населяют весьма разнообразные условия наземной среды и поведение их также весьма разнообразно. Они встречаются в песчаных, глинистых и каменистых пустынях, в зарослях густой травы, в лесу, на болотах и т. д. Вместе с тем большинство предпочитает открытые участки, где много солнца и растительность не мешает передвижению.
Характер передвижения рептилий весьма разнообразен, и только некоторые из них могут быть названы «пресмыкающимися» в полном смысле этого термина, т. е. животными, волочащими свое тело по земле. Крокодилы, вараны и многие ящерицы бегают, высоко приподняв тело на ногах. Некоторые сухопутные черепахи также не волочат брюхо по земле. Есть виды, способные пробегать большое расстояние на одних только задних ногах. Таковы некоторые агамы, игуаны и особо характерная в этом отношении австралийская плащеносная ящерица (Chlamydosaurus). Особенно много таких рептилий было среди вымерших мезозойских групп.
Способность пустынных рептилий бегать, высоко приподнимая свое тело на ногах, несомненно, имеет приспособительное значение, так как при таком положении тело не испытывает губительного воздействия раскаленного солнцем грунта. Виды рептилий, живущие в песчаных пустынях, обладают рядом при-способлений для передвижения по сыпучему грунту. Чаще всего они выра-жаются в наличии на краях пальцев оторочки из роговых зубчиков. Эти приспособления свойственны видам различных систематических группЗ агамы, гекконы, ящерицы, игуаны. У одного из видов гекконов (Palmatogecko rangi), живущего в песчаных пустынях Южной Африки, между пальцами имеются кожистые перепонки.
Некоторые рептилии песчаных пустынь имеют уплощенное, широкое тело. Так, у одного из видов игуан (Phrynosoma), живущего в Мексике, форма тела имеет вид диска. Замечательно, что подобная же приспособительная к жизни на сыпучем песке черта свойственна и нашей среднеазиатской круглоловке (Phrynocephalus).
Обширная и разнообразная по составу группа рептилий приспособлена к передвижению в густой траве, среди кустов и в кучах растительного хлама. В основе эта приспособленность заключается в удлинении тела, получающего «змеевидную» форму, и в частичной или полной редукции конечностей. Наиболее совершенно эта черта организации развита у змей, большинство видов которых полностью утеряли не только свободные конечности, но и их пояса. Рудименты задних конечностей имеются у удавов, слепунов (Typhlopidae), у коралловой змеи (Ilysia). Но и среди ящериц есть полностью безногие виды. Такова, например, наша веретеница. У желтопуза есть едва заметные рудименты задних конечностей. Есть, наконец, виды с очень слаборазвитыми конечностями, которые при быстром передвижении в большей мере пользуются змеевидными изгибами удлиненного тела, чем конечностями. Это свойственно многим сцинкам: гологлазу (Ablepharus,), халциду (Chalcides).
Механизм движения рептилий со змеевидным телом весьма разнообразен. Большинство змей изгибают тело в горизонтальной плоскости, но некоторые ползают с прямо вытянутым телом. Движение в этом случае осуществляется за счет последовательного перемещения кожи брюха сзади наперед и обратно.
Очень многие рептилии хотя бы часть времени проводят под землей, в которую закапываются сами, или залезают в норы других животных, в трещины в почве. Приспособления к рытью разнообразны. Сухопутные черепахи роют норы обеими парами конечностей; в норах они спасаются от дневной жары. Так же поступают и некоторые ящерицы. Но большинство, закапываясь, действуют головой, с помощью которой раздвигают грунт. Межчелюстной щиток многих роющихся змей выдается вперед и служит как бы заступом. Из наших змей легко зарывается в песок и свободно ползает под его покровом песчаный удав (Erix). Рептилии, ведущие подземный образ жизни, имеют удлиненное, змеевидное тело и полностью или частично редуцированные конечности. Чешуя у них гладкая или редуцированная. Слабо развиты или атрофированы глаза. В качестве примера укажем на слепунов (Typhlops), амфисбен (Amphisbaenidae).
Весьма своеобразно закапываются в песке круглоголовки. По бокам их уплощенного тела располагаются кожистые оторочки, снабженные че-шуйками. Спасаясь от врагов, животное плотно прижимается к земле и, быстро перемещая тело то вправо, то влево, разгребает песок, который засыпает его сверху. Животное мгновенно утопает в песке, опускаясь в него не под углом (как это делают змеи и другие ящерицы), а отвесно вниз. Подобные приспособления имеются и у некоторых змей (рогатая гадюка, эфа).
Среди рептилий есть много видов, хорошо лазающих по отвесным плитам камней, кустам и деревьям. Эта особенность свойственна различным систематическим группам и связана с разнообразными приспособлениями.
У некоторых ящериц сильно развиты пальцы с длинными изогнутыми когтями и длинный неотламывающийся хвост, который может закручиваться вокруг ветвей и служит дополнительной точкой опоры. Наиболее совершенное приспособление такого рода свойственно хамелеонам. У них пальцы передних и задних ног срастаются в две противоположные друг другу группы, и животное обхватывает ими ветки, как клещами. Среди наших ящериц хорошо лазают по кустам агамы. Есть лазающие виды и среди змей.
Немногим видам свойственна способность к планирующему полету, развившаяся в связи с древесным образом жизни. У живущего в лесах Зондских островов лопатохвостого геккона (Ptychozoon) по бокам головы, тела, хвоста и между пальцами имеется кожистая перепонка, служащая парашютом. Еще более совершенное приспособление свойственно летающим драконам (Draco) — обитателям лесов Малайского архипелага. Парашютом служит широкая складка кожи, находящаяся по бокам тела и при прыжке растяги¬вающаяся 5—6 парами ребер. Длина прыжка у этого вида достигает 20 м. Эта ящерица нередко на лету ловит насекомых.
Значительное число рептилий ведут в той или иной мере водный образ жизни. Как и в предшествующих случаях, приспособленность к водному существованию наблюдается среди различных по систематическому положению рептилий. Различен и характер приспособлений. Крокодилы плавают при помощи сжатого с боков хвоста; лишь отчасти плаванию у них помогают лапы. Морская игуана (Amblyrhynchus), живущая на Галапагосских островах, плавает только при помощи хвоста, а лапы держит плотно прижатыми к туловищу. Изгибая хвостовую часть тела (также сжатую с боков), плавают морские змеи.
<
Совершенно иное приспособление имеют водные черепахи; орудием плавания у них служат ластообразные конечности.
Наряду с орудиями плавания у водных рептилий имеются и другие приспособительные особенности. Напомним о клапанах, закрывающих ноздри. У настоящих водных черепах в той или иной мере редуцируется костный и роговой панцирь. Изменены органы дыхания. Легкие имеют сложное строение и обеспечивают лучший газообмен при больших интервалах между вдохами. Кроме того, у черепах и водных змей в глотке появляются вздутые участки, богатые кровеносными сосудами и являющиеся своего рода внутренними «жабрами».
Питание
Рептилии питаются самыми различными животными — от мелких наземных и водных беспозвоночных до крупных млекопитающих. Ряд видов едят растительную пищу, но все же большинство добывают животные корма. Так, ящерицы чаще ловят наземных насекомых, червей и моллюсков. Кроме того, некоторые из них (агамы, игуаны) охотно поедают и растительную пищу. Морские черепахи и морские змеи обычно пита¬ются рыбами. Ужи истребляют много амфибий, а некоторые морские змеи, например пеламида (Pelamydrus), кормятся почти исключительно головоногими моллюсками. Крупные ящерицы, например вараны, и многие наземные змеи добывают мелких зверьков, птиц и пресмыкающихся. Крокодилы и крупные змеи едят не только мелких животных, но и крупных зверей, например молодых оленей. Известны случаи нападения их на людей.
Настоящими растительноядными рептилиями являются наземные черепахи (например, степная черепаха). Лишь иногда они поедают мелких животных. Большинство пьют воду, и только виды, обитающие в пустынях, получают воду из съеденной пищи.
Способы отыскивания и добывания пищи различны. Ящерицы и некоторые змеи много бродяжничают и разыскивают пищу. Добычу они ловят как на поверхности, так и под землей. По наблюдениям Н. В. Шибанова, сетчатая ящурка (Eremias grammica) в пустынях Средней Азии выкапывает из песка насекомых, зарывающихся на глубину до 10 см. Сцинки, роющиеся в песке на глубине нескольких сантиметров, легко обнаруживают пищу, находящуюся на поверхности, и как бы выныривают из песка, чтобы схватить ее. Некоторые змеи (например, удавы), крокодилы и водные черепахи подкарауливают добычу и бросаются на нее. Караулят, сидя на ветвях, свою добычу и хамелеоны.
Почти все рептилии глотают пищу целиком, не разделяя ее на части и не подвергая ее во рту измельчению. Только черепахи и крокодилы отделяют куски пищи. Надо вспомнить, что именно у этих видов имеется вторичное костное нёбо, позволяющее задерживать пищевой комок во рту без помехи для дыхания. Большинство рептилий пожирают свою добычу, не умерщвляя ее предварительно. Только ядовитые змеи и удавы глотают уже убитую жертву.
Размножение
Размножение рептилий существенно отличается от размножения амфибий и имеет много приспособительных к наземному существованию особенностей. Оплодотворение всегда внутреннее, и самцы имеют копулятивные органы. Яйца развиваются вне воды, в тех или иных условиях наземной среды. Развитие проходит без превращения, т. е. в отличие от амфибий личиночной стадии не бывает, и вылупившаяся из яйца молодь живет в тех же условиях, что и взрослые.
Основные приспособления к размножению вне воды, на суше, заключаются в особенностях строения и развития яйца. Сравнительно с амфибиями рептилии имеют крупные яйца. Укрупнение яиц идет за счет обогащения их желтком, а у некоторых (черепах и крокодилов) и за счет появления «белка». Большой запас питательного вещества в яйце обусловливает возможность прямого (без превращения) развития эмбрионов. В связи с откладыванием яиц на суше возникает ряд приспособлений, защищающих их от иссушения, механического повреждения и обеспечивающих для развивающегося эмбриона возможность газообмена, снабжения его водой и выведения наружу (хотя бы частично) продуктов распада. Приспособления эти выражаются в основном в появлении ряда оболочек, покрывающих яйцо.
Яйца всех рептилий покрыты прочной волокнистой оболочкой, предохраняющей их от растекания, механических повреждений и от проникновения бактерий; волокнистая оболочка защищает яйцо и от высыхания.
У черепах и крокодилов, кроме того, имеется белковая оболочка (подобная белку птичьего яйца), которая является основным запасом воды, обеспечивающим развитие зародыша. У прочих рептилий необходимая для нормального развития яйца вода имеет двоякую природу. Часть ее возникает в результате окисления жира (весьма обильного в желтке) — так называемая метаболическая вода, часть же поступает из наружной среды через оболочки яйца.
Укажем для сравнения, что, в то время как у амфибий процент жира в яйце равен всего 8—11, у рептилий он достигает 40 и более.
Кроме описанных волокнистой, известковой и белковой оболочек, продуцируемых соответствующими отделами яйцеводов, в развивающемся яйце рептилий (как и у прочих амниота) по мере хода развития, за счет самого яйца, возникает еще ряд оболочек, имеющих исключительно важное приспособительное значение; таковы амнион, сероза и аллантоис.
Еще на ранних стадиях развития зародыш погружается в желток. Впереди от головного отдела зародыша возникает складка, известная под названием амниотической, которая разрастается назад и в итоге замыкает зародыш. Возникающий таким путем амнион представляет сплошной замкнутый мешок, внутри которого находится зародыш. Полость амниона (амниотическая полость) заполняется жидкостью. Таким образом, зародыш находится как бы в маленьком водоеме, условия среды которого относительно постоянны. Роль амниона заключается еще и в том, что он предохраняет зародыш от соприкосновения с твердыми яйцевыми оболочками.
Если учесть наличие ряда оболочек, отгораживающих зародыш от наружной среды, то, естественно, возникает вопрос о том, как дышит зародыш и как выводятся наружу продукты обмена. Первоначальным приспособлением для дыхания служит густая сеть кровеносных сосудов, оплетающих желток, — желточный круг кровообращения. В дальнейшем таковой постепенно замещается новым образованием — сетью сосудов, развивающихся в аллантоисе. Аллантоис возникает как мешковидный вырост брюшной стенки задней кишки; по мере развития аллантоис приобретает вид большого пузыря, лежащего между желточным мешком и серозой, с которой срастается передняя часть аллантоиса. К концу развития аллантоис полностью одевает тело зародыша и желточный мешок.
Аллантоис выполняет одновременно две функции: он является органом дыхания и зародышевым мочевым пузырем. При этом аллантоис не только является пассивным приемником мочи, но восринимает из мочи излишнюю воду, которая через сеть кровеносных сосудов вводится в тело, вновь выделяется с мочой и т. д. Кроме того, часть продуктов обмена зародыша выводится наружу в газообразном состоянии. Это обусловливается тем, что основным энергетическим материалом в яйце рептилий является жир, окисляющийся до воды и СO2.
Все описанные особенности строения и развития яиц рептилий носят ясно выраженный приспособительный характер. Они:
1) предохраняют яйца от растекания, механических повреждений, иссушения (волокнистая и известковая оболочки);
2) обеспечивают возможность прямого развития (обилие питательного материала — желтка);
3) обеспечивают поступление воды в яйцо из наружной среды (гигроскопичность яйца и пористость оболочек);
4) обеспечивают запас воды в самом яйце (жир желтка, а у некоторых — богатый водой белок яйца);
5) создают относительно постоянные условия для эмбриона (жидкость амниотической полости);
6) обеспечивают возможность дыхания и выведения газообразных продуктов обмена в яйце, покрытом рядом оболочек (аллантоис).
Эти принципиальные особенности размножения имели решающее значение при переходе рептилий к наземному обитанию.
Большинство рептилий размножаются путем откладывания оплодотворенных яиц во внешнюю среду, обычно в специально сооружаемую в земле ямку, в сравнительно влажных и хорошо обогреваемых солнцем местах. Есть виды, откладывающие яйца в кучи растительного мусора, в гнилые пни. Американские крокодилы, вырыв в болотистой почве ямку, засыпают яйца гниющими растениями.
Как правило, рептилии, отложив яйца, покидают их. Но некоторые крокодилы остаются у «гнезда» и охраняют яйца от других животных. Самки питонов обвиваются вокруг яиц своим тело, обеспечивая таким путем не только сохранность яиц, но и относительно постоянную температуру вокруг них. В гнезде питонов она на 2—4° С выше температуры окружающей среды. Охраняет яйца и самка варана.
Наряду с откладыванием яиц у некоторых рептилий наблюдается яйцеживорождение и живорождение. В простейшем случае оплодотворенные яйца остаются в половых путях и проходят здесь все стадии развития. Детеныши вылупляются из яиц тотчас же после откладывания их во внешнюю среду. Так бывает у гадюки, живородящей ящерицы (Lacerta vivipara) и веретеницы (Anguis fragilis).
Любопытно, что у ужей (Natrix natrix) срок развития яиц вне тела матери варьирует от 30 до 60 суток, что связано с большей или меньшей продолжительностью периода нахождения оплодотворенных яиц в теле матери. В отложенных яйцах зародыши оказываются сформированными примерно наполовину. Очень большая разница в длине периода внутриутробного нахождения яиц наблюдалась и у нашей прыткой ящерицы (15—20 суток), при этом зародыши оказываются развитыми на 1/3 и более. Таким образом, предпосылкой для развития яйцеживорождения служит возможность более или менее длительной задержки оплодотворенных яиц в теле матери. Яйцеживорождение свойственно, кроме указанных видов, некоторым удавам, в частности-нашему песчаному удавчику (Eryx), морским змеям, многим ужам, ящерицам,. Ни у одного из видов черепах и крокодилов яйцеживорождение не наблюдается.
Для немногих видов свойственно настоящее живорождение, при котором кровеносные сосуды желточного мешка тесно сближены с сосудами маточного отдела яйцевода и питание эмбриона идет в значительной степени за счет организма матери. Таковы некоторые сцинки.
Наконец, у некоторых змей и ящериц формируется настоящая аллантоидная плацента, при которой ворсинки хориоаллантоиса внедряются в слизистую половых путей (Вauсhоt, 1965; Hoffman, 1970).
В явлении живородности (включая сюда и яйцеживородность) есть довольно ясно выраженные закономерности. Установлено, что наибольший процент живородных видов свойствен северным широтам и высокогорным областям. К этому надо добавить, что обнаружены виды, которые в разных частях ареалов бывают то яйцеродными, то живородными. Так, тибетская круглоголовка (Phrynocephalus theobaldi) на высоте 2—3 тыс. м размножается откладыванием яиц, а на высоте 4—5 тыс. м она живородна. Установлено, что издавна считавшаяся живородящей ящерица Lacerta viviрага на крайнем юге своего ареала (Франция) откладывает яйца.
Очевидно, что основной причиной, вызывающей живородность у рептилий, является холодный климат. При нахождении яиц в теле матери обеспечивается более или менее постоянная и высокая температура, необходимая для их успешного развития. В отличие от млекопитающих, имеющих постоянную температуру тела, у рептилий это достигается тем, что самка своим поведением, перемещаясь в наиболее хорошо обогреваемые места, регулирует термические условия развития зародышей. Дополнительной причиной живородности, проявляющейся у немногих видов, является водный и отчасти древесный и подземный образ жизни. Так, живородящи морские змеи и некоторые хамелеоны.
Количество яиц, откладываемых рептилиями, сравнительно невелико. В редком случае оно достигает 50—100 или несколько более. Так бывает у некоторых крокодилов, у крупных черепах и змей. Обычно число яиц не превышает 20—30. Мелкие виды ящериц откладывают всего 1—2 яйца. Любопытно, что в этом случае бывает несколько кладок в год. Откладывание яиц в несколько приемов свойственно и многим черепахам, и в частности видам черепах, которые водятся в России. В общем плодовитость рептилий заметно меньшая, чем амфибий и тем более рыб. Это стоит в прямой связи с более совершенной организацией всего процесса размножения рептилий.
Половая зрелость наступает в различные сроки. Так, у крокодилов и большинства черепах это бывает в 8—10-летнем возрасте, у змей — чаще в возрасте 3—5 лет, у ящериц — обычно на 2—3-м году жизни, но некоторые мелкие виды становятся половозрелыми уже на следующую весну, т. е. в возрасте 9—10 месяцев. Крайне интересно, что у некоторых скальных ящериц, распространенных в Восточном Закавказье (например, Lacerta armenica), самцы неизвестны вовсе или встречаются крайне редко. Самки у них размножаются партеногенетически и рождают только самок. Лишь немногие виды соединяются в пары на длительное время. Таковы некоторые агамы, ящерицы и немногие черепахи. Большинство расходятся после совокупления. По некоторым наблюдениям (Caulfield, 1966), самки миссисипского аллигатора не только строят гнездо и охраняют его, но и помогают выведшимся молодым выбраться из гнезда. Молодые держатся вместе с матерью по крайней мере в течение одного сезона.
Поделиться с друзьями