
Происхождение и эволюция рептилий
Происхождение рептилий
Первые позвоночные появились на суше в девоне. Это были стегоцефалы, или панцирноголовые амфибии,— ближайшие родственники кистеперых рыб. Подобно последним, они значительную часть времени проводили в водоемах. Однако при периодически повторяющихся засухах они могли выползать из пересыхающих водоемов и в поисках более благоприятных условий некоторое время проводить на суше.
Происхождение рептилий. Возможность все более и более долгое время оставаться на суше определялась благоприятными условиями последующего каменноугольного периода: климат был влажный, теплый и ровный на большей части, видимо, единого материка. Но уже в конце каменноугольного периода условия существования на суше изменились. Громадные горообразовательные процессы, перемещения участков суши относительно полюсов Земли вызывали перемену климатов и растительности. Во многих областях Земли климат стал засушливым, континентальным. Годичные кольца на стволах деревьев указывают на разницу условий существования по сезонам года. Зимы, видимо, были холодными. Пышная растительность из хвощей и папоротников, связанная с озерами и болотами, исчезла. Появились обширные пустынные пространства. Все большее преобладание получала относительно сухолюбивая растительность из хвойных и саговников.
Условия существования для стегоцефалов стали неблагоприятными. Сухость воздушной среды затрудняла длительное пребывание их на поверхности земли, так как легочное дыхание у них было несовершенным, а голая кожа не могла препятствовать иссушению тела. В то же время пустынный во многих областях ландшафт не обеспечивал возможности размножения стегоцефалов, которые откладывали свои яйца в воду. Большинство стегоцефалов вымерло еще до наступления пермского периода. Но одновременно указанные условия среды вызывали появление у наиболее сухопутных из них ряда новых приспособительных признаков.
Решающими приспособлениями, обеспечивающими возможность жить полностью на суше, были:
- прогрессивное развитие центральной нервной системы, обеспечивающей более совершенное приспособительное поведение животных;
- ороговение верхнего слоя эпидермиса, а затем и появление роговых чешуи, что предохраняло тело от иссушения;
- увеличение количества желтка в яйце и появление при его развитии ряда оболочек, защищающих зародыш от иссушения и вместе с тем обеспечивающих возможность газообмена.
Животные получили возможность жить и размножаться на суше. Естественно, что одновременно появились и другие особенности организма. Усилились конечности, скелет стал более прочным. Усложнились легкие, ставшие теперь единственным органом дыхания.
Эволюция рептилий
Эволюция рептилий шла очень быстро и бурно. Еще задолго до конца пермского периода они вытеснили большинство стегоцефалов. Получив возможность существовать на суше, рептилии в новой среде столкнулись с новыми и крайне разнообразными ее условиями. Воздействие таких разнообразных условий жизни и отсутствие на суше существенной конкуренции со стороны других животных и послужили той основной причиной, которая обусловила чрезвычайно бурный расцвет рептилий в последующее время. Они получили возможность и вместе с тем были вынуждены приспособиться к самым различным условиям наземной среды. В последующем многие из них вторично в той или иной мере приспособились к жизни в воде. Некоторые стали воздушными животными. Приспособительное расхождение рептилий было поразительным. С полным основанием мезозой считают веком рептилий.
Первичные рептилии
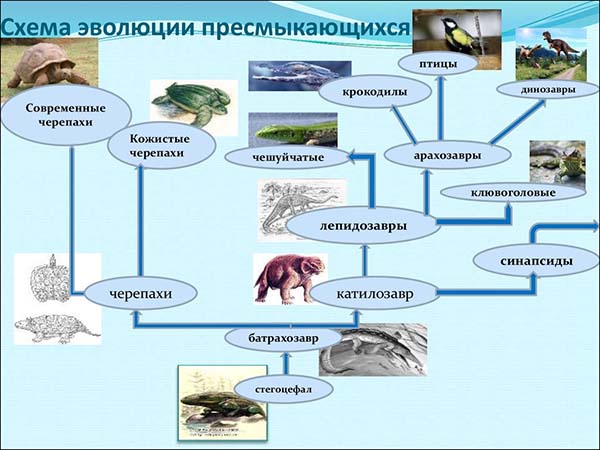
Котилозавры — древнейшие рептилии, известные из верхних каменноугольных отложений.
По ряду признаков они еще очень близки к стегоцефалам. Так, у многих был только один крестцовый позвонок; шейный отдел развит слабо, в плечевом поясе имелся клейтрум — кожная кость, свойственная рыбам. Череп был в виде сплошной костной коробки с отверстиями только для глаз, ноздрей и теменного органа (отсюда название этой группы — цельночерепные). Конечности были коротки и не специализированы.
Среди в общем немногочисленных котилозавров наиболее примитивным будут сеймурия (Seymouria), обнаруженная в пермских отложениях Северной Америки, и близкие к ней формы, найденные на Северной Двине, также в пермских отложениях. Это были некрупные животные, размером не более 0,5 м. Больших размеров достигали парейазавры (Pareiasaurus), многочисленные остатки которых найдены В. П. Амалицкимна Северной Двине. Размеры их достигали 3 м. Большинство котилозавров были растительноядными, некоторые питались моллюсками.
Котилозавры достигали расцвета в средней перми. Но лишь немногие сохранились до конца перми, а в триасе эта группа исчезла, уступив место более высокоорганизованным и специализированным группам рептилий, развившимся из различных отрядов котилозавров.
Дальнейшая эволюция рептилий обусловливалась изменчивостью их в связи с воздействием весьма разнообразных условий жизни, с которыми они сталкивались при размножении и расселении. Большинство групп приобрели большую подвижность; скелет их стал более легким, но в то же время и более прочным. Рептилии использовали все более и более разнообразную пищу. Изменялась техника ее добывания. В этой связи существенным изменениям подверглось строение конечностей, осевого скелета и черепа. У большинства конечности стали длиннее, таз прикреплялся к двум и более крестцовым позвонкам. В плечевом поясе исчезла кость клейтрум. Сплошной панцирь черепа подвергся частичной редукции. В связи с более дифференцированной мускулатурой челюстного аппарата в височной области черепа возникли ямы и разделяющие их костные мосты — дуги, служившие для прикрепления сложной системы мускулов.
Ниже рассматриваются главнейшие группы рептилий, обзор которых должен показать исключительное разнообразие этих животных, их приспособительную специализацию и вероятное соотношение с ныне живущими группами.
Первоящеры (Prosauria) — одна из наиболее примитивных групп рептилий, череп которых имел две скуловые дуги. Зубы, как и у амфибий, сидели не только на костях челюстей, но и на нёбе. Позвонки были амфицельные, как и у рыбы и низших амфибий. Внешностью похожи на крупных ящериц. Наиболее древние представители известны из пермских отложений. В триасе появляются представители хоботноголовых (Rhynchocephalia), один из видов которых — гаттерия (Sphenodon punctatus) — сохранился до настоящего времени в Новой Зеландии.
Псевдозухии (Pseudosuchia) произошли, вероятно, от одного корня с первоящерами. Впервые появляются в начале триаса. По общему облику и размерам отчасти были похожи на ящериц. Своеобразные черты организации заключались в том, что зубы сидели в глубоких ячейках; задние конечности были развиты значительно сильнее передних, и у большинства только они и использовались для хождения. В этой связи таз и нижние отделы скелета задних конечностей были удлинены. Многие вели, по-видимому, древесный образ жизни. Таков, например, орнитозух.
Псевдозухии, несомненно, близки к крокодилам, птерозаврам и динозаврам, для развития которых они, видимо, и служили исходной группой. Наконец, есть основание считать, что псевдозухии дали начало и предкам птиц.
Крокодилы (Crocodilia) появляются в конце триаса. Юрские крокодилы существенно отличны от современных отсутствием настоящего костного нёба, и внутренние ноздри у них открывались между нёбными костями. Позвонки были еще амфицельными. В меловой период существовали крокодилы современного типа с полностью развитым вторичным костным нёбом и продельными позвонками. Большинство обитало в пресных водоемах, но среди юрских форм известны и настоящие морские виды.
Крылатые ящеры (Pterosauria) представляют один из замечательных примеров специализации мезозойских рептилий. Это были летающие животные весьма своеобразного строения. Орудием полета служили крылья, представлявшие складку кожи, натянутую между боками тела и очень длинным четвертым пальцем передних конечностей. Широкая грудина имела хорошо развитый киль, как и у птиц, кости черепа срастались рано, многие кости были пневматичными. Вытянутые в клюв челюсти имели у некоторых видов зубы. Длина хвоста и форма крыльев варьировали. У одних (рамфоринхи) крылья были длинные, узкие и длинный хвост; летали они, видимо, скользящим полетом, часто планируя. У других (птеродактили) хвост был очень коротким, а крылья широкие; полет их чаще был гребной. Судя по тому, что остатки птерозавров обнаружены в отложениях соленых водоемов, это были обитатели побережий. Они питались рыбой и по поведению, видимо, были близки к чайкам и крачкам. Размеры варьировали от нескольких сантиметров до метра и более. Наибольшего расцвета птерозавры достигли в юре. Отдельные виды известны и из меловых отложений.
Динозавры (Dinosauria) — следующая, последняя ветвь псевдозухий, виды которой жили с начала триаса до конца мела. Это наиболее многочисленная и разнообразная группа рептилий. Среди динозавров были мелкие животные с длиной тела менее метра и гиганты почти до 30 м длины. Одни из них ходили только на задних ногах, другие на всех четырех ногах. Очень разнообразен был и общий внешний облик тела, но у всех голова была относительно мала, а спинной мозг в крестцовой области образовывал местное расширение, объем которого превосходил объем головного мозга.
Динозавры в самом начале их обособления от псевдозухий разделились на две ветви, развитие которых шло параллельно. Характерной особенностью их были черты строения тазового пояса, в связи с чем эти группы именуются птицетазовые и ящеротазовые.
Ящеротазовые первоначально были сравнительно мелкими хищными животными, передвигавшимися скачками только на задних ногах, в то время как передние служили для схватывания пищи. Для опоры служил и длинный хвост. В последующем появились крупные растительноядные формы, ходившие на всех четырех ногах. К ним относятся самые крупные позвоночные, когда-либо жившие на суше. Так, бронтозавр имел длину тела около 20 м, а диплодок — до 26 м. Большинство гигантских ящеротазовых, видимо, были полуводными животными и питались сочной водной растительностью.
<
Птицетазовые получили свое название в связи с удлиненным тазом, похожим на таз птиц. Первоначально они передвигались на одних удлиненных задних ногах, но более поздние виды имели соразмерно развитые обе пары конечностей и ходили на четырех ногах. По характеру питания птицетазовые были исключительно растительноядными животными. Среди них упомянем игуанодонов, ходивших только на задних ногах и достигавших 9 м высоты. Кожа их была без костного панциря. Трицератопс внешне был очень похож на носорога, обладал обычно небольшим рогом на конце морды и двумя длинными рогами над глазами. Длина его доходила до 8 м. Стегозавр был характерен несоразмерно маленькой головой и двумя рядами высоких костных пластинок, располагавшихся на спине. Длина его была равна около 5 м.
Динозавры были распространены почти по всему земному шару и обитали в крайне разнообразных жизненных условиях. Они населяли пустыни, леса, болота. Некоторые (например, траходонты) вели полуводный образ жизни. Несомненно, что в мезозое динозавры были господствующей на суше группой рептилий. Они появились в триасе и наибольшего расцвета достигли в меловое время. К концу этого периода динозавры вымерли.
Чешуйчатые (Squamata). История этого самого многочисленного в настоящее время отряда наименее ясна.
Ящерицы появились, видимо, еще в верхней юре, но только в меловой период наблюдается относительное разнообразие этого подотряда. Змеи развились позже всех остальных рептилий. Они появились лишь к концу мела, несомненно, как боковой ствол ящериц. Настоящий расцвет чешуйчатых наступил только в третичное время, когда вымерло большинство групп рептилий.
Черепахи (Chelonia) представляют одну из древнейших труп рептилий, происшедших, видимо, непосредственно от котилозавров. Предком их принято считать пермского Eunotosaurus. Это небольшое ящерообразное животное с короткими и очень широкими ребрами, образующими подобие спинного щита. Брюшного щита у них не было. Имелись зубы. В триасе появляются уже настоящие черепахи с развитым настоящим панцирем (например, Triassochelys).
Однако голова и конечности у них еще не могли полностью втягиваться в панцирь. Был развит роговой чехол на челюстях, но одновременно на нёбе имелись зубы. Мезозойские черепахи первоначально были сухопутными и, видимо, роющими животными. Лишь впоследствии некоторые группы перешли к водному образу жизни и в этой связи частично утратили костный и роговой панцирь.
За все время с триаса и до наших дней черепахи сохранили все основные черты своей организации. Они пережили все испытания, которые погубили большинство рептилий, и в настоящее время процветают в такой же мере, как и в мезозое.
Ихтиозавры (Ichthyosauria)—рептилии, наиболее полно приспособившиеся к жизни в воде. В природе мезозоя они занимали то же место, которое ныне занимают китообразные. Их конвергентное сходство с дельфинами поразительно. Они имели веретенообразное тело, вытянутое рыло и большой двухлопастный плавник. Парные конечности были превращены в ласты, при этом задние конечности и таз недоразвились. Фаланги пальцев были удлинены, а число пальцев у некоторых доходило до 8. Кожа была голой. Размеры тела варьировали от 1 до 14 м. Ихтиозавры жили только в воде и питались рыбой, отчасти беспозвоночными. Установлено, что они были живородящи. Появление ихтиозавров относится к триасу. В меловой период они вымерли. Генетические соотношения с другими рептилиями не выяснены.
Плезиозавры (Plesiosauria) — вторая группа мезозойских морских рептилий с иными приспособительными особенностями организации. Ихтиозавры плавали, волнообразно изгибая тело и особенно хвостовую его часть, плавники у них служили для управления. Плезиозавры имели широкое и плоское тело со сравнительно слаборазвитым хвостом. Орудием плавания служили мощные ласты. В отличие от ихтиозавров у них была хорошо развита шея, несущая небольшую голову. Размеры тела от 50 см до 15 м. Отличен был, видимо, и образ жизни. Во всяком случае некоторые виды населяли прибрежные воды. Питались рыбой и моллюсками.
Плезиозавры появились в начале триаса. В конце мелового периода они вымерли.
Звероподобные (Theromorpha) представляют очень большой интерес как группа, давшая начало млекопитающим.
Звероподобные — одна из наиболее древних групп рептилий. Появление ее относится к концу каменноугольного времени, и в перми они были уже многочисленны и разнообразны. Период своего расцвета звероподобные пережили еще задолго до того, как появились первые динозавры, и котилозавры были их непосредственными родичами. Примитивные звероподобные, выделяемые в отряд Пеликозавров (Pelycosauria), были еще очень близки к котилозаврам. Так, они имели двояковогнутые позвонки и хорошо сохранившиеся брюшные ребра. Однако зубы их сидели в альвеолах, а в височной области черепа имелась боковая впадина, не свойственная ни одной другой группе рептилий. Внешностью они походили на ящериц и имели небольшие размеры — 1—2 м. У некоторых намечалась, хотя и в небольшой степени, дифференцировка зубов (например, у Sphenacodon).
В средней перми пеликозавров сменили более высокоорганизованные зверозубые (Theriodontia). Зубы у них были явственно дифференцированы, появилось вторичное костное нёбо. Единый затылочный мыщелок разделился на два. Нижняя челюсть в основном была представлена зубной костью. Положение конечностей также изменилось. Локоть переместился назад, а колено вперед, и в итоге конечности начали занимать положение под туловищем, а не по бокам его, как у других рептилий. В скелете появилось много черт, общих с млекопитающими.
Среди многочисленных пермских звероподобных были весьма разнообразные по облику и образу жизни рептилии. Многие были хищниками. Такова, например, иностранцевия (Inostrancevia aiexandrovi), найденная экспедицией В. П. Амалицкого в отложениях пермского периода на Северной Двине. Другие питались растительной или смешанной пищей. Эти малоспециализированные виды и стоят ближе всего к млекопитающим. Среди них надо указать на циногнатуса (Cynognathus), имевшего много прогрессивных черт организации.
Зверозубые были многочисленные еще в триасе, но при появлении хищных динозавров они исчезли.
Из приведенного выше обзора филогении рептилий видно, что подавляющее большинство крупных систематических групп их (отрядов) вымерли до начала кайнозойской эры, и современные рептилии представляют лишь жалкие остатки мезозойской фауны.
Причина этого грандиозного явления понятна лишь в самых общих чертах. Обращает внимание, что большинство мезозойских рептилий представляли собой чрезвычайно специализированных животных. Успех их существования зависел от наличия весьма своеобразных, узкоопределенных жизненных условий. Надо думать, что односторонняя специализация была одной из предпосылок исчезновения большинства мезозойских рептилий.
Установлено, что, хотя вымирание отдельных групп рептилий наблюдалось в течение всего мезозоя и конца палеозоя, особенно резко оно было выражено в конце мезозоя, именно в конце мелового периода. В это время за сравнительно короткий срок вымерло огромное большинство мезозойских пресмыкающихся. Если справедливо наименование мезозоя как века рептилий, то не менее оправдано назвать конец этой эры веком великого вымирания. Наряду со сказанным установлено, что в меловое время наблюдались особо значительные изменения климатов и ландшафтов. Это было связано с существенными перераспределениями суши и моря и движениями земной коры, приведшими к громадным горообразовательным явлениям, известным в геологии под названием «альпийской стадии горообразования». Нарушения сложившихся условий жизни в этой связи были весьма значительными. Они заключаются не только в изменении климатов, орографии Земли и других условий мертвой природы. Достаточно указать, что в середине мелового периода мезозойская флора хвойных, саговниковых и других сменилась растениями нового типа, именно покрытосеменными. Естественно, что все это не могло не отразиться на успехе существования всех животных, и однобоко специализированных в первую очередь.
Наконец, надо учесть, что к концу мезозоя все большее и большее развитие получали несравненно более высокоорганизованные — птицы и млекопитающие, сыгравшие очень большую роль в борьбе за существование меж¬ду группами наземных животных.
Поделиться с друзьями