
Экология рыб
Некоторые условия жизни рыб в водной среде
Вода как жизненная среда обладает рядом специфических особенностей, создающих своеобразные условия существования.
Жизненная арена рыб исключительно велика. При общей поверхности земного шара, равной примерно 510 млн. кв. км, около 361 млн. кв. км, т. е. 71% всей площади, занято поверхностью океанов и морей. Кроме того, около 2,5 млн. кв. км, или 0,5% площади земного шара, занято внутренними водоемами. Обширность жизненной арены определяется, кроме того, и большим простиранием ее по вертикали. Максимальная известная глубина океана равна примерно 11 тыс. м. Океаны с глубиной более 3 тыс. м занимают примерно 51—58% от всей площади морских вод. Далее следует учесть, что рыбы живут в областях, расположенных от экватора до приполюсовых пространств; они есть в горных водоемах на высоте более 6 тыс. и над уровнем моря и в океанах на глубине более 10 тыс. м. Все это создает большое разнообразие условий существования. Разберем некоторые из черт водной среды обитания применительно к населяющим ее рыбам.
Подвижность водной среды связана с постоянными течениями в реках и морях, местными течениями в мелких замкнутых водоемах, вертикальными перемещениями слоев воды, обусловленными различным их прогреванием.
Подвижность воды обусловливает в значительной мере пассивные перемещения рыб. Так, личинки норвежской сельди, которые вывелись у берегов Западной Скандинавии, увлекаются одной из ветвей течения Гольфстрим на северо-восток и за 3 месяца относятся вдоль побережья на 1000 км.
Мальки многих лососевых рыб выводятся в вершинах притоков крупных рек, а основную часть жизни они проводят в морях. Переход из рек в моря совершается также в значительной мере пассивно; они сносятся в моря течениями рек.
Наконец, подвижность воды обусловливает пассивные передвижения кормовых объектов —планктона, что в свою очередь влияет на перемещение рыб.
Колебания температуры в водной среде значительно меньшие, чем в среде воздушно-наземной. В подавляющем большинстве случаев верхний предел температуры, при которой встречаются рыбы, лежит ниже +30, +40° С. Особенно же характерен нижний предел температуры воды, который даже в сильно соленых частях океанов не опускается ниже —2° С. Следовательно, реальная амплитуда температур среды обитания рыб равняется всего 35—45° С.
Вместе с тем надо учесть, что и эти сравнительно ограниченные колебания температур имеют в жизни рыб большое значение. Влияние температуры осуществляется как прямым воздействием на организм рыб, так и косвенным, через изменение способности воды растворять газы.
Как известно, рыбы принадлежат к так называемым холоднокровным животным. Температура их тела не остается более или менее постоянной, как у теплокровных животных, — она находится в прямой зависимости от температуры окружающей среды. Это связано с физиологическими особенностями организмов, в частности с характером процесса теплообразования. У рыб этот процесс идет значительно медленнее. Так, карп массой 105 г выделяет в сутки на 1 кг массы 42,5 кдж тепла, а скворец массой 74 г на 1 кг массы выделяет в сутки 1125 кдж. Известно, что температура среды, а следовательно, и температура тела рыб существенно влияют на такие важные биологические явления, как созревание половых продуктов, развитие икры, питание. Понижение температуры воды вызывает у ряда рыб спячку. Таковы, например, карась, сазан, осетровые.
Косвенное влияние температуры воды хорошо может быть прослежено на особенностях явлений газообмена у рыб. Известно, что способность воды растворять газы, и в частности кислород, обратно пропорциональна ее температуре и солености.
Вместе с тем потребность рыб в кислороде увеличивается по мере повышения температуры воды. В связи с изложенным изменяется и минимальная концентрация кислорода, ниже которой рыба погибает. Для карпа она будет равна: при температуре 1° С — 0,8 мг/л, при температуре 30° С — 1,3 мг/л, а при 40°С — около 2,0 мг/л.
В заключение укажем, что потребность различных видов рыб в кислороде неодинакова. По этому признаку они могут быть разбиты на четыре группы: 1) требующие очень много кислорода; нормальные условия для них — 7—11 см3 кислорода на литр: кумжа (Salmo trutta), гольян (Phoxinus phoxinus), голец (Nemachilus barbatulus); 2) требующие много кислорода — 5—7 см3 на литр: хариус (Thymallus thymallus), голавль (Leuciscus cephalus), пескарь (Gobio gobio); 3) потребляющие сравнительно небольшое количество кислорода — около 4 см3 на литр: плотва (Rutilus rutilus), окунь (Perea fluviatilis), ерш (Acerina cernua); 4) выдерживающие очень слабое насыщение воды кислородом и живущие даже при 1/2 см3 кислорода на литр: сазан, линь, карась.
Образование льда в водоемах имеет в жизни рыб огромное значение. Покров льда в известной мере изолирует нижележащие слои воды от низких температур воздуха и тем самым предотвращает промерзание водоема до дна. Это делает возможным распространение рыб в областях с очень низкими температурами воздуха зимой. Таково положительное значение ледяного покрова.
Покров льда играет в жизни рыб и отрицательную роль. Это сказывается в его затемняющем действии, которое замедляет или даже почти вовсе приостанавливает жизненные процессы у многих водных организмов, прямо или косвенно имеющих для рыб пищевое значение. В первую очередь это касается зеленых водорослей и высших растений, которыми кормятся частью сами рыбы и те беспозвоночные животные, которых поедают рыбы.
Ледяной покров чрезвычайно резко снижает возможность пополнения воды кислородом из воздуха. Во многих водоемах зимой в результате гнилостных процессов совершенно теряется кислород, растворенный в воде. Возникает явление, известное под названием замора водоемов. У нас в стране оно имеет распространение и наблюдается в бассейнах, водосборная площадь которых связана в значительной мере с болотами (чаще торфяными). Большие заморы наблюдались в бассейне Оби. Болотные воды, питающие здесь реки, богаты гуминовыми кислотами и закисными соединениями железа. Эти последние, окисляясь, отнимают из воды растворенный в ней кислород. Возмещение же его из воздуха невозможно в связи со сплошным покровом льда.
Из рек огромной территории Западной Сибири рыбы уже в декабре начинают спускаться в Обь и, следуя по ней вниз, достигают в марте Обской губы. Весной, по мере таяния льда, рыба поднимается обратно (так называемый вонзевой ход рыбы). Заморы наблюдаются и в Европейской части России. С заморами ведут успешную борьбу путем сооружения прорубей или путем увеличения проточности пруда или озера. В прудовых хозяйствах, имеющих высокую техническую оснащенность, употребляются компрессоры, нагнетающие воду с кислородом. На подходе рыб к прорубям или к специально сооруженным в берегах озера отепленным канавам основан один из способов лова рыбы. Любопытно, что поселение на некоторых подвергающихся замору водоемах бобров и ондатр ослабило это явление, так как через норы, хаты и другие сооружения этих зверей облегчается газообмен между водоемами и атмосферой.
Звукопроводность воды очень велика. Это обстоятельство широко используется рыбами, среди которых широко развита звуковая сигнализация. Она обеспечивает получение информации как среди особей одного вида, так и сигналы о наличии особей других видов. Не исключено, что издаваемые рыбами звуки имеют эхолокационное значение.
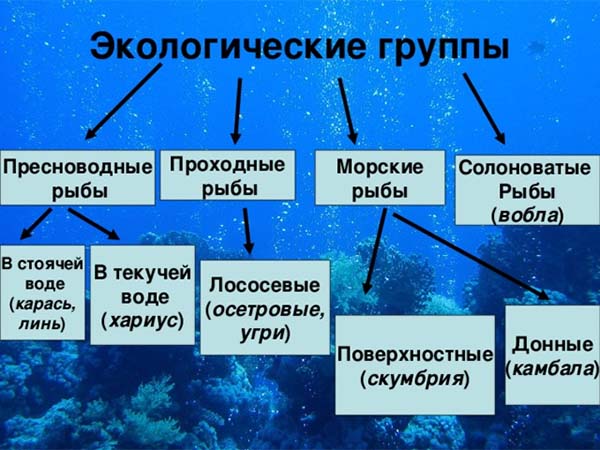
Экологические группы рыб
Морские рыбы
Это наиболее многочисленная группа видов, которые всю жизнь проводят в соленой морской воде. Населяют разнообразные горизонты, и по этому признаку следует различать такие группы.
1. Пелагические рыбы. Обитают в толще воды, в которой широко перемещаются в поисках пищи и мест, пригодных для размножения. Подавляющее большинство плавает активно и имеет удлиненное, веретенообразное тело; таковы, например, акулы, сардины, макрели. Немногие, например луна-рыба, перемещаются в значительной мере пассивно с токами воды.
2. Литорально-придонные рыбы. Обитают в придонных слоях воды или на дне. Здесь они находят корм, нерестятся и спасаются от преследования. Распространены на различных глубинах, от мелководья (скаты, некоторые камбалы, бычки) до значительных глубин (химеровые).
Способность к плаванию худшая, чем у видов предыдущей группы. Многие имеют разнообразные приспособления для пассивной защиты в виде шипов, колючек (некоторые скаты, бычки), толстого наружного панциря (кузовок).
3. Абиссальные рыбы. Немногочисленная Группа, населяющая глубоко-водные (ниже 200 м) части морей и океанов. Условия их существования крайне своеобразны и в общем неблагоприятны. Это обусловливается отсутствием на больших глубинах света, низкими температурами (не выше +4° С, чаще около 0° С), огромным давлением, большей соленостью воды, отсутствием растительных организмов. Абиссальные рыбы частью лишены глаз, частью же, наоборот, обладают огромными телескопическими глазами; некоторые имеют органы свечения, облегчающие поиски пищи. В связи с отсутствием растений все абиссальные рыбы плотоядны; они или хищники, или падалееды.
<
Пресноводные рыбы
Пресноводные рыбы обитают только в пресных водоемах, из которых не выходят даже в осолоненные предустьевые участки морей. В зависимости от типа водоема среди пресноводных рыб различают следующие группы:
1. Рыбы стоячих вод обитают в озерах и прудах (карась, лини, некоторые сиги).
2. Общепресноводные рыбы населяют стоячие и текучие воды (щука, окунь).
3. Рыбы текучих вод. В качестве примера можно указать на форель, жереха.
Проходные рыбы
Проходные рыбы в зависимости от стадии жизненного цикла обитают то в морях, то в реках. Почти все проходные рыбы проводят период роста и созревания половых продуктов в море, а для икрометания идут в реки. Таковы многие лососевые (кета, горбуша, семга), осетровые (осетр, белуга), некоторые сельди. В качестве противоположного примера надо указать на речных угрей (европейского и американского), которые размножаются в море (Атлантический океан), а период подготовки к нересту проводят в реках.
Рыбы этой группы зачастую совершают очень длительные миграции протяженностью в 1000 и более километров. Так, кета из северной части Тихого океана заходит в Амур, по которому поднимается (некоторые косяки) выше Хабаровска. Европейский угорь из рек Северной Европы идет на икрометание в Саргассово море, т. е. западную часть Атлантического океана.
Полупроходные рыбы
Полупроходные рыбы обитают в предустьевых опресненных частях морей, а для размножения, а в некоторых случаях и для зимовок заходят в реки. Однако в отличие от настоящих проходных рыб они не поднимаются высоко по рекам. Таковы вобла, лещ, сазан, сом. Эти рыбы местами могут жить и оседло в пресных водоемах. Группа полупроходных рыб наименее естественная.
Форма тела некоторых групп рыб
В связи с исключительным разнообразием условий обитания внешний вид рыб также крайне разнообразен. Большинство видов, населяющих открытые пространства водоемов, имеют веретенообразное тело, часто несколько сжатое с боков. Это хорошие пловцы, так как быстрота плавания в указанных условиях необходима и для хищных рыб при ловле добычи, и для мирных рыб, вынужденных спасаться от многочисленных хищников. Таковы акулы, лососи, сельди. Основным органом поступательного движения у них служит хвостовой плавник.
Среди рыб, обитающих в открытых частях водоемов, сравнительно немногочисленны так называемые планктонные рыбы. Они живут в толще воды, но передвигаются часто пассивно вместе с течениями. Внешне большинство из них отличается укороченным, но сильно расширенным телом, иногда почти шарообразной формы. Плавники развиты очень слабо. Примером могут служить еж-рыба (Diodon) и меланоцетус (Melanocetus). У луны-рыбы (Mola mola) тело очень высокое, сжатое с боков. Хвостового и брюшного плавников у нее нет. Иглобрюх (Spheroides) после наполнения кишечника воздухом становится почти шарообразным и плавает по течению брюхом вверх.
Значительно более многочисленны и разнообразны придонные рыбы. Глубоководные виды имеют часто каплеобразную форму, при которой рыба обладает большой головой и постепенно утончающимся к хвосту туловищем. Таковы длиннохвост (Macrurus norvegicus) и химера (Chimaera monstrosa) из хрящевых рыб. Близки по форме тела к ним треска и бельдюга, живущие в придонных слоях, иногда на значительных глубинах. Вторым типом придонных глубоководных рыб являются уплощенные в спинно-брюшном направлении скаты и уплощенные с боков камбалы. Это малоподвижные рыбы, которые кормятся также медленно передвигающимися животными. Среди донных рыб встречаются виды, имеющие змеевидное тело, — угри, морские иглы, вьюны. Они обитают среди зарослей водной растительности, и передвижение их похоже на движение змей. Наконец, упомянем о своеобразных кузовках (Ostracion), тело которых заключено в костный панцирь, предохраняющий рыбу от пагубного действия прибоя.
Жизненный цикл рыб, миграции
Как и всем живым существам, рыбам на разных этапах их жизненного пути необходимы различные условия среды. Так, условия, необходимые для нереста, отличны от условий, обеспечивающих наилучший нагул рыб, своеобразные условия нужны для зимовки и т. д. Все это приводит к тому, что в поисках условий, подходящих для каждого данного жизненного отправления, рыбы совершают более или менее значительные перемещения. У видов, населяющих мелкие замкнутые водоемы (пруды, озера) или реки, перемещения имеют ничтожные масштабы, хотя и в этом случае они все же выражены вполне отчетливо. У морских и особенно у проходных рыб миграции развиты наиболее сильно.
Наиболее сложны и разнообразны нерестовые миграции у проходных рыб; они связаны с переходом из морей в реки (чаще) или, наоборот, из рек в моря (реже).
Переход для размножения из морей в реки (анадромные миграции) свойствен многим лососевым, осетровым, некоторым сельдям и карповым. Значительно меньше видов, кормящихся в реках и для нереста идущих в моря. Такие перемещения называют катадромными миграциями. Они свойственны угрям. Наконец, и многие чисто морские рыбы совершают в связи с нерестом длительные перемещения, переходя из открытого моря к берегам или, наоборот, от прибрежий в глубину моря. Таковы морские сельди, треска, пикша и др.
Протяженность пути нерестовых миграций весьма различна в зависимости от вида рыб и условий населяемых ими водоемов. Так, виды полупроходных карповых северной части Каспия поднимаются вверх по рекам всего на несколько десятков километров.
Громадные по протяженности миграции совершают многие лососевые. У дальневосточного лосося — кеты — миграционный путь достигает местами двух и более тысяч километров, а у нерки (Oncorhynchus nerka) — около 4 тыс. км.
Семга поднимается по Печоре до ее верховьев. Несколько тысяч километров проходит на пути к местам нереста европейский речной угорь, размножающийся в западной части Атлантического океана.
Длина миграционного пути зависит от того, насколько приспособлены рыбы к условиям, в которых может осуществляться нерест, а в указанной связи и от того, как далеко от мест нагула расположены места, подходящие для нереста.
Время нерестовых миграций у рыб вообще не может быть указано столь же определенно, как, например, сроки миграций птиц на гнездовье. Это связано, во-первых, с тем, что сроки нереста у рыб весьма разнообразны. Во- вторых, известно много случаев, когда рыбы подходят к местам нереста почти за полгода до самого нереста. Так, например, семга Белого моря входит в реки в два срока. Осенью идут особи со сравнительно слаборазвитыми половыми продуктами. Они зимуют в реке и размножаются на следующий год. Наряду с этим есть и другая биологическая раса беломорской семги, которая входит в реки летом, — половые продукты у этих особей хорошо развиты, и они мечут икру в том же году. У кеты также бывает два нерестовых хода. В Амур «летняя» кета заходит в июне — июле, «осенняя» — в августе — сентябре. В отличие от семги обе биологические расы кеты нерестуют в год захода в реку. Вобла заходит в реки для нереста весной, некоторые сиги, наоборот, мигрируют к местам размножения только осенью.
Приведем обобщенные описания нерестовых миграций некоторых видов рыб.
Морская норвежская сельдь перед размножением нагуливается далеко к северо-западу от Скандинавии, у Фарерских островов, и даже в водах у Шпицбергена. В конце зимы косяки сельди начинают двигаться к берегам Норвегии, которых они достигают в феврале—марте. Нерест происходит в фиордах близ берега на мелких местах. Тяжелая икра, выметанная рыбами, в огромном количестве оседает на дно и приклеивается к водорослям и камням. Выведшиеся личинки лишь отчасти остаются в фиордах; большая же их масса увлекается Нордкапским течением (северо-восточная ветвь Гольфстрима) вдоль берегов Скандинавии на север. Такую пассивную миграцию личинки начинают зачастую в еще очень раннем возрасте, когда у них сохраняется желточный пузырь. За три-четыре месяца, до конца июля — начала августа, они проделывают путь в 1000 — 1200 км и достигают берегов Финмаркена.
Обратный путь молодые сельди проходят активно, но значительно более медленно—за четыре-пять лет. Они продвигаются на юг этапами ежегодно, то приближаясь к берегам, то отходя в открытое море. В четырех- или пятилетием возрасте сельдь становится половозрелой, и к этому времени она достигает района икрометания — места, где она родилась. Этим заканчивается первый, «юношеский» этап ее жизни — период далекого путешествия на север.
Второй период, период зрелости, связан с ежегодными миграциями от места нагула к местам нереста и обратно.
Рассмотрим на примере дальневосточных лососевых нерестовые миграции проходных рыб. Жизнь их в море в период полового созревания недостаточно изучена. Однако все же известно, что, вопреки распространенному ранее мнению, лососи держатся не распыленно по огромной акватории севера Тихого океана, а концентрируются в определенных его районах.
При миграциях на нерест скорость движения рыб, насколько это удалось выяснить выловом меченых особей, весьма значительна. Так, в одном случае из трех меченых горбуш одна прошла за 33 суток 900 км, другая — 1200 км и третья — более чем 1400 км, идя с рекордной быстротой по 44 км в сутки. Кета, помеченная в Тихом океане под 45°00' с. ш. и 148°10/ в. д., была поймана на северном берегу Охотского моря, в Тауйской губе, т. е. более чем в 1600 км от места выпуска.
Ход рыбы в реке лучше всего изучен на Амуре В. К. Солдатовым и его последователями. Как уже указано, там наблюдаются два ее хода: летний — с конца июня и осенний — в августе — сентябре.
Кета двигается вверх по течению со значительной скоростью — 30 — 35 км, а по некоторым данным, даже и 47 и в сутки. Если к этому учесть, что Амур течет со средней скоростью в 68 км, то фактически рыба проходит в сутки до 115 км. Заходя в притоки Амура и двигаясь вверх по мелким речкам, рыба вынуждена преодолевать перекаты и пороги, где струи воды бьют с очень большой силой. Водопады до 1 м высоты рыба перепрыгивает вполне успешно. Перед препятствием она обычно отдыхает и набирается сил в месте со спокойной водой; затем быстро устремляется к водопаду и делает прыжок, вылетая из воды. Прыгая, рыба очень часто попадает на гребень водопада и сносится вниз. Отдохнув, она вновь повторяет свою попытку.
Преодолевая препятствия, рыба, естественно, расходует огромное количество энергии. По П. Ю. Шмидту, расход энергии в сутки у кеты равен для самцов 103 240 кдж, для самок — 113 560 кдж на каждый килограмм живой массы. При этом надо учесть, что, заходя в реки, рыбы перестают питаться и расход энергии осуществляется целиком за счет накопленных в теле питательных веществ. Все сказанное приводит к резким изменениям внешнего вида и физиологического состояния мигрантов.
По данным П. Ю. Шмидта, при движении от моря до нерестилища, находящегося на расстоянии 1200 км, кета теряет жира: самец — 98,7%, самка — 97,3%; белка: самец — 57,3%, самка — 57,7%; золы: самец — 47,0%, самка — 47,1%; воды: самец — 15,2%, самка — 20,7%.
По мере движения к местам нереста происходит созревание половых продуктов. Так, у кеты, идущей в районе Амурского лимана, масса гонад самки составляет 7% от общей массы рыбы, на расстоянии 300 км от устья масса гонад увеличивается до 10% от массы тела, на расстоянии 570 км —до 12%, на расстоянии 1000 км — до 17% и у места нереста (на расстоянии 1190 км) — до 19%.
Из моря кета выходит стройной серебристой рыбой, но уже в низовьях реки серебристый блеск чешуи исчезает и окраска сильно темнеет. По бокам появляются поперечные лиловые, а затем и малиновые полосы, разделенные яркими черными полосками. Меняется и форма тела. Оно становится выше и площе с боков, на спине возникает горб. Конец рыла загибается крючком вниз в виде клюва, а нижняя челюсть на переднем конце загибается вверх. Чем дальше поднимается рыба, тем сильнее видны эти изменения. Она покрывается ранами, язвами, обрастает грибком сапролегнией. К местам нереста кета приходит в состоянии крайнего истощения.
Места нереста расположены в притоках Амура, большей частью в их верховьях. Обычно это участки со спокойным течением, с галечным грунтом. Глубина воды 0,5—1,2 м. Кета появляется здесь с развитыми половыми продуктами и вскоре же начинает нереститься. Икру кета откладывает в специально вырытые ямки и засыпает галькой.
Проделав огромный и трудный путь, израсходовав почти всю энергию на дорогу и самый процесс размножения, рыбы, которые за весь этот период ничего не едят, вскоре же после нереста погибают. Одни погибают уже на местах нереста, другие сносятся течением рек вниз и погибают на обратном пути в море. Берега рек бывают усеяны трупами кеты, большое количество зверей и птиц скапливается здесь и уничтожает ослабевшую или уже погибшую рыбу. Несомненно, что возвращений в море у кеты не бывает и вся масса зашедших для нереста рыб погибает. Таким образом, кета размножается один раз в жизни. Мальки кеты весной следующего года начинают двигаться вниз по течению и летом достигают моря. Здесь они становятся половозрелыми и через 3—5 лет отправляются в странствование к местам, где они родились, но откуда они живыми уже не вернутся.
Чрезвычайно сложны и интересны связанные с размножением миграции речного угря. Вид этот распространен по берегам Европы и Северной Африки, где обитает в самых разнообразных пресных водоемах: в реках, ручьях, прудах. Способность угрей расселяться очень велика, так как они могут переползать по суше из одного водоема в другой. Такие сухопутные миграции совершаются ночью, угри ползут по влажной траве иногда несколько километров.
Жизнь угрей в реках издавна была хорошо известна. Она начинается с появлением в устьях рек небольших угревидных рыбок длиной 6—8 см и с совершенно прозрачным телом. Поднимаясь по рекам, угри темнеют и при длине около 20 см тело их покрывается чешуей. Продолжительность жизни их в реках различна, но во всяком случае не менее 6, а часто 10 и более лет.
Достигнув определенного возраста (в разных условиях различного), угри приобретают брачный наряд и начинают мигрировать вниз по рекам в моря. Это явление внешне малозаметно, так как угри идут чаще ночью и по глубоким местам рек. По выходе в море угри становятся почти недоступными для наблюдений, и долгое время не было известно значение этих миграций. Загадкой был и характер размножения угрей, так как у особей, живущих в реках, половые продукты незаметны. Это породило самые фантастические мнения. Так, Аристотель полагал, что угри возникают в иле в болотистых озерах. В последующем высказывалось мнение о живородности угрей, основанное на нахождении в их теле паразитических червей, которых считали за развивающуюся молодь. Только в конце XIX в. путем гистологических исследований удалось установить наличие семенников и яичников. Стало очевидным, что угри размножаются обычным половым путем. Но еще долгое время оставалось неясным, где же проходит их нерест. Ученые не знали ни зрелой икры угрей, ни их молоди.
Наряду со сказанным еще с начала прошлого столетия рыбакам и зоологам была известна маленькая прозрачная рыбка столь своеобразного строения, что ее выделили в особый род — Leptocephalus. Лептоцефалы отличались различными размерами и строением; их было описано несколько видов. Только в 1897 г. опытным путем удалось установить, что лептоцефалы представляют личинок угрей, и проследить их превращение в тех прозрачных угрей, заходы которых из морей в реки наблюдались издавна. Однако вопрос о месте размножения угрей оставался невыясненным.
Решить эту загадку удалось знаменитому датскому ихтиологу Иоганну Шмидту, который начиная с 1905 г. предпринимал систематические исследования в морях, омывающих Западную Европу, и в Атлантическом океане. Оказалось, что в восточных окраинных морях Атлантического океана (Северном и Балтийском) встречаются либо прозрачные угри, либо лептоцефалы, проходящие метаморфоз. По мере движения к юго-западу в Атлантическом океане встречаются лептоцефалы все меньших размеров. Самые мелкие лептоцефалы, длиной менее 10 мм, были обнаружены только в одной области, расположенной между Бермудскими и Багамскими островами. Здесь они держатся на глубине 200—300 м.
Жизненный цикл угрей оказался расшифрованным, и в схеме он представляется так. Взрослые угри выходят из рек Европы и Северной Африки и, двигаясь на запад или юго-запад, достигают западных частей Атлантического океана. Они двигаются со средней скоростью 15 км в сутки, но иногда скорость достигает 40—50 км в сутки. Их путь равен 7—8 тыс. км. Весной угри опускаются на глубину около 1000 м, нерестятся и погибают. Выведшиеся личинки поднимаются из мрачных глубин, куда не проникает ни один луч света, на залитую тропическим солнцем поверхность океана и начинают обратное странствование на восток через весь океан. Они двигаются в значительной мере пассивно, влекомые струями Гольфстрима. Три года продолжается это путешествие, и вот у берегов Европы и Африки появляются прозрачные угри, которые входят в реки, где начинают вторую фазу своей жизни. Через 6—9 лет (а иногда и через 10—20 лет), уже взрослыми, они вновь направляются в океан, где оставляют потомство, а сами погибают. Ни один угорь не возвращается в реку.
Чем ориентируются рыбы, отыскивая места нереста, отстоящие от мест нагула на тысячи километров? Как и в связи с чем возникли явления нерестовых миграций?
В общем виде дать ответ на первый вопрос сравнительно легко.
Несомненно, что в основе миграционных движений рыб лежит их нервнорефлекторная деятельность. При миграциях рыбы испытывают закономерные и постепенные изменения условий среды, играющие роль своего рода сигналов — раздражителей.
Так, проходные лососи, видимо, ориентируются постепенными изменениями химизма воды, наблюдаемыми по мере движения из открытых пространств моря к предустьевым областям и далее к самим рекам. Меняется соленость воды, богатство ее кислородом, щелочность или содержание в ней углекислоты. Мигрирующие угри руководствуются, видимо, в основном закономерными изменениями температурных условий и солености воды. Дело в том, что область их нереста является самой теплой областью в океане и областью максимальной солености. Таким образом, угри, вышедшие из любого пункта европейского или американского побережья, если они будут ориентироваться постоянно повышающейся температурой воды и увеличением ее солености, должны будут прийти кратчайшим путем к области наиболее теплой и соленой воды, т. е. к месту своего икрометания.
Несомненно, что не зрение, а удивительная по тонкости способность воспринимать особенности химического и физического состояния воды и связанная с этим феноменальная «память» на соответствующие раздражения «руководит» рыбами при их тысячеверстных странствованиях. При этих миграциях рыба «устремляется к источнику максимального раздражения, совершенно так же, как ночная бабочка летит на свет фонаря, раздражающий ее орган зрения» (П. Ю. Шмидт, 1947).
Однако все сказанное не объясняет, почему рыбы идут на нерест, например, из Тихого океана в Амур или из рек Европы в центральные части Атлантики.
Миграции угрей еще не получили должного объяснения. Согласно данным П. Ю. Шмидта, во время максимального оледенения распределение температур в Атлантике было иным и область наиболее теплой воды располагалась далеко к югу от современного ее положения и имела вид не овала ограниченных размеров в западных частях океана, как в настоящее время, а составляла более или менее широкую полосу, проходившую через весь океан с запада на восток. Миграционный путь угрей, как европейских, так и американских, был в этой связи коротким. С окончанием ледникового периода, когда Гольфстрим принял свое современное северо-восточное направление, а с другой стороны, в восточную часть океана проникли холодные воды южного полушария, область максимальных температур сократилась и отодвинулась к западу. Угри по мере отодвигания ледника и появления на материках новых рек заселяли их. Расселение угрей на север и сокращение и отодвигание области наиболее теплой воды на запад и вызвали удлинение миграционных путей.
На миграции проходных лососей существуют два противоположных взгляда. То обстоятельство, что почти все проходные рыбы распространены в северном полушарии, делает весьма вероятным зависимость возникновения миграций от явлений ледникового периода. По одному из предположений, опреснение морских вод в период таяния ледников сделало возможным выход взрослых речных рыб в моря, более богатые кормом, чем реки. В реки рыбы возвращались для размножения, так как условия для этого здесь более благоприятны: меньше хищников и конкурентов, лучше химические особенности воды, например обеспеченность кислородом и др.
[sch:11]
По другой гипотезе, проходные рыбы были исконно морскими и заход их в реки — явление вторичное, связанное с сильным опреснением морей во время таяния ледников, что в свою очередь позволило рыбам легче приспосабливаться к жизни в пресной воде. Так или иначе, но несомненно, что проходные лососевые меняют места своего обитания в зависимости от особенностей биологического состояния. Взрослые рыбы населяют обширные пространства морей, богатые кормом. Их молодь выводится в тесных пресных водоемах (верховья рек), где невозможно было бы существование всей массы выросших рыб в силу ограниченности самого пространства и из-за недостатка корма. Однако для вывода молоди условия здесь более благоприятны, чем в море. Это обусловливается чистой, богатой кислородом водой, возможностью закапывания икры в грунт дна и возможностью ее успешного развития в пористом грунте. Все это настолько благоприятствует успеху размножения, что количество икринок, обеспечивающее сохранение вида, доходит, например, у горбуши всего до 1100—1800 икринок.
Кормовые миграции в тех или иных масштабах свойственны практически всем рыбам. Естественно, что в мелких замкнутых водоемах перемещения рыб в поисках пищи носят весьма ограниченный характер и внешне резко отличаются от длительных и массовых странствований, наблюдаемых у морских или проходных рыб.
Природа кормовых миграций в общем смысле вполне понятна, если учесть, что в период нереста рыбы выбирают весьма специфические условия среды, которые, как правило, не представляют большой ценности в кормовом отношении. Вспомним, например, что лососевые и осетровые мечут икру в реках с их весьма ограниченными для громадных масс зашедших рыб пищевыми возможностями. Уже одно это обстоятельство должно вызвать перемещение рыб после нереста. Кроме того, большинство рыб во время размножения перестают питаться, и, следовательно, после нереста потребность в корме резко возрастает. В свою очередь сказанное заставляет рыб искать области с особо благоприятными кормовыми возможностями, что и усиливает их перемещения. Примеров кормовых миграций очень много среди разнообразных биологических групп рыб.
Европейский лосось — семга, в отличие от своего тихоокеанского сородича — кеты, не погибает полностью после нереста, и передвижения отнерестившихся рыб вниз по реке должны рассматриваться как кормовые миграции. Но и после выхода рыб в море они совершают массовые закономерные миграции в поисках мест, особо богатых пищей.
Так, каспийская севрюга, вышедшая после нереста из Куры, пересекает Каспийское море и нагуливается преимущественно у восточного побережья Каспия. Молодь кеты, скатившаяся на следующую (после нереста) весну вниз по Амуру, идет на откорм к берегам Японских островов.
Не только проходные, но и морские рыбы показывают примеры отчетливо выраженных кормовых миграций. Норвежская сельдь, нерестующая на отмелях у юго-западного побережья Скандинавии, не остается после размножения на месте, а массами перемещается на север и северо-запад, к Фарерским островам и даже в Гренландское море. Здесь, на границе теплых вод Гольфстрима и холодных вод арктического бассейна, развивается особо богатый планктон, на котором и откармливаются истощенные рыбы. Любопытно, что одновременно с миграцией на север сельди в этом же на-правлении мигрирует и сельдевая акула (Lanina cornubica).
Широко мигрирует в поисках пищи атлантическая треска. Одним из основных мест ее нереста служат отмели (банки) у Лофотенских островов. После размножения треска становится чрезвычайно прожорливой, и в поисках пищи большие ее стаи направляются частью вдоль берегов Скандинавии на северо-восток и далее на восток через Баренцево море к острову Колгуеву и Новой Земле, частью на север, к Медвежьему острову и далее к Шпицбергену. Эта миграция представляет для нас особый интерес, так как лов трески в районе Мурманска и в Канинско-Колгуевском мелководье в значительной мере базируется на добыче мигрирующих и кормящихся косяков. Треска при миграциях придерживается теплых струй Нордкапского течения, по которым, согласно новейшим данным, проникает через Карские Ворота и Югорский Шар даже в Карское море. Наибольшее количество трески в Баренцевом море скапливается в августе, но уже с сентября начинается ее обратное движение, и к концу ноября крупная, пришлая от берегов Норвегии треска в наших водах исчезает. К этому времени температура воды резко падает и становится неблагоприятной как для самих рыб, так и для животных, служащих им пищей. Треска, напитавшись и накопив жир в печени, начинает обратное движение на юго-запад, ориентируясь температурой воды, служащей хорошим ориентиром — раздражителем при миграциях.
Протяженность пути, совершаемого в один конец треской при описанных миграциях, равна 1—2 тыс. км. Рыбы двигаются со скоростью 4—11 морских миль в сутки.
Наряду с горизонтальными миграциями известны случаи вертикальных перемещений морских рыб в поисках пищи. Скумбрия поднимается в поверхностные слои воды тогда, когда здесь наблюдается наиболее богатое развитие планктона. При опускании планктона в более глубокие слои туда же опускается и скумбрия.
Зимовальные миграции. Многие виды рыб при зимнем понижении температуры воды становятся малоактивными или даже впадают в состояние оцепенения. В этом случае они не остаются обычно в местах нагула, а собираются на ограниченных пространствах, где условия рельефа, дна, грунта и температуры благоприятствуют зимовке. Так, сазан, лещ, судак мигрируют в низовья Волги, Урала, Куры и других больших рек, где, скапливаясь в огромном количестве, залегают в ямы. Издавна известна зимовка осетров в ямах на реке Урал. Наши тихоокеанские камбалы в летнее время распространены по всему заливу Петра Великого, где не образуют больших скоплений. Осенью, по мере понижения температуры воды, рыбы эти отходят от берегов в глубины и собираются в немногих местах.
Физической причиной, вызывающей у рыб своего рода спячку, служит понижение температуры воды. В состоянии спячки рыбы лежат неподвижно на дне, чаще в углублениях дна — ямах, где скапливаются нередко в огромном количестве. У многих видов поверхность тела в это время покрывается толстым слоем слизи, которая в известной мере изолирует рыб от отрицательного действия низких температур. Обмен веществ у зимующих таким образом рыб чрезвычайно сильно понижается. Некоторые рыбы, например караси, зимуют, закапываясь в ил. Известны случаи, когда они вмерзают в ил и успешно перезимовывают, если «соки» их тела не подвергаются замораживанию. Опыты показали, что лед может окружать все тело рыбы, но внутренние «соки» остаются незамерзшими и имеют температуру до -0,2,-0,3° С.
Не всегда зимовальные миграции заканчиваются впадением рыб в состояние оцепенения. Так, азовская хамса по окончании нагула на зиму выходит из Азовского моря в Черное. Это связано, видимо, с неблагоприятными температурными и кислородными условиями, возникающими зимой в Азовском море в связи с появлением ледяного покрова и сильным охлаждением воды этого неглубокого водоема.
Ряд приведенных выше примеров показывает, что жизненный цикл рыб складывается из ряда последовательно сменяющих друг друга этапов: созревания, размножения, нагула, зимовки. Во время каждого из этапов жизненного цикла рыбы нуждаются в различных специфических условиях среды, которые они находят в различных, зачастую далеко отстоящих друг от друга местах водоема, а иногда и в разных водоемах. Степень развитости миграции неодинакова у разных видов рыб. Наибольшее развитие миграции получают у проходных рыб и рыб, живущих в открытых морях. Это понятно, так как разнообразие условий среды обитания в этом случае очень велико и в процессе эволюции у рыб могло выработаться важное биологическое приспособление — существенно менять места обитания в зависимости от этапа биологического цикла. Естественно, что у рыб, населяющих небольшие и особенно замкнутые водоемы, миграции меньше развиты, что соответствует и меньшему разнообразию условий в таких водоемах.
Характер жизненного цикла у рыб различен и по другим признакам.
Некоторые рыбы, и их большинство, ежегодно (или через некоторые интервалы) нерестятся, повторяя одни и те же передвижения. Другие в течение жизненного цикла только один раз проходят стадию созревания половых продуктов, один раз предпринимают нерестовую миграцию и только однажды в жизни размножаются. Таковы некоторые виды лососевых (кета, горбуша), речные угри.
Питание
Характер пищи у рыб чрезвычайно разнообразен. Рыбы кормятся почти всеми живыми существами, обитающими в воде: от мельчайших планктонных растительных и животных организмов до крупных позвоночных. При этом сравнительно немногие виды кормятся только растительной пищей, большинство же поедает животные организмы или смешанную животно-растительную пищу. Деление рыб на хищных и мирных в значительной мере условно, так как характер пищи существенно варьирует в зависимости от условий водоема, времени года и возраста рыбы.
Особо специализированными растительноядными видами являются планктона явные толстолобы (Hyspophthalmichthys) и поедатели высшей растительности белые амуры (Ctenopharyngodon).
Из рыб нашей фауны преимущественно растительные виды следующие: красноперка (Scardinius), маринка (Schizothorax) и храмуля (Varicorhinus). Большинство рыб кормится смешанной пищей. Однако в молодом возрасте все рыбы проходят стадию мирного питания планктоном и лишь в последующем переходят на свойственную им пищу (бентос, нектон, планктон). У хищников переход на рыбный стол происходит в различном возрасте. Так, щука начинает заглатывать личинок рыб, достигнув длины тела всего 25—33 мм, судак — 33—35 мм; окунь же переходит на рыбное питание сравнительно поздно, при длине тела в 50—150 мм, при этом беспозвоночные все же составяют главную пищу окуня в течение 2—3 первых лет его жизни.
В связи с характером питания устройство ротового аппарата у рыб существенно различно. У хищных видов рот вооружен острыми загнутыми назад зубами, которые сидят на челюстях (а у рыб с костным скелетом нередко также и на нёбных костях и на сошнике). Скаты и химеры, питающиеся донными беспозвоночными, одетыми панцирем или скорлупой, имеют зубы в виде широких плоских пластинок. У рыб, грызущих кораллы, зубы имеют вид резцов и нередко срастаются в одно целое, образуя острый режущий клюв. Таковы зубы сростночелюстных (Plectognathi).
Кроме настоящих челюстных зубов, у некоторых рыб развиваются еще так называемые глоточные зубы, которые сидят на внутренних краях жаберных дуг. У карповых рыб они расположены на нижнем отделе задней видоизмененной жаберной дуги и получают название нижнеглоточных зубов. Эти зубы перетирают пищу о роговую мозолистую площадку, расположенную на нижней стороне мозгового черепа, — так называемый жерновок. У губанов (Labridae) имеются верхние и нижние глоточные зубы, расположенные друг против друга; жерновок в этом случае отсутствует. При наличии глоточных зубов настоящие челюстные зубы или отсутствуют вовсе, или развиты слабо и лишь помогают схватыванию и удерживанию пищи.
Приспособление к роду пищи видно не только в строении зубов, но и в устройстве всего ротового аппарата. Выделяют несколько типов ротового аппарата, главнейшие из которых следующие:
1. Хватательный рот широкий, с острыми зубами на челюстных костях, а часто и на сошнике и нёбных костях. Жаберные тычинки в этом случае коротки и служат для предохранения жаберных лепестков, а не для процеживания пищи. Характерен для хищных рыб: щуки, судака, сома и многих других.
2. Рот планктонояда средней величины, обычно не выдвижной; зубы малы или отсутствуют. Жаберные тычинки длинные, действующие, как сито. Свойственен сельдям, сигам, некоторым карповым.
3. Рот всасывательный имеет вид более или менее длинной трубки, иногда выдвигающейся. Работает, как всасывающая пипетка, при питании донными беспозвоночными или мелкими планктонными организмами. Таков рот леща, морской иглы. Особенное развитие этот тип ротового аппарата получил у африканских длиннорылов (Mormyridae), которые в поисках пищи засовывают свое трубкообразное рыло под камни или в ил.
4. Рот бентосоеда — скатов, камбал, осетровых — расположен на нижней стороне головы, что связано с добычей пищи со дна. В некоторых случаях рот вооружен мощными жерновообразными зубами, служащими для раздробления панцирей и раковин.
5. Рот с ударными или мечевидными челюстями или рылом. В этом случае челюсти (сарганы — Belonidae) или рыло (скаты, пила-рыба — Pristis, акулы-пилоносы — Pristiophorus) сильно вытянуты и служат для нападения на стаи рыб, например сельдей. Есть и другие типы ротового аппарата, полный перечень которых нет нужды здесь приводить. Отметим в заключение, что даже у систематически близких рыб легко можно видеть отличия в устройстве рта, связанные с характером питания. Примером могут служить карповые рыбы, питающиеся то донными, то планктонными, то падающими на поверхность воды животными.
Кишечный тракт тоже существенно варьирует в зависимости от характера питания. У хищных рыб, как правило, кишечник короткий, а желудок хорошо развит. У рыб, кормящихся смешанной или растительной пищей, кишечник значительно длиннее, а желудок слабо обособлен или совсем отсутствует. Если в первом случае кишечник лишь несколько превосходит длину тела, то у некоторых растительноядных видов, например у закаспийской храмули (Varicorhinus), он в 7 раз длиннее тела, а у толпыги (Hypophthalmichthys), питающейся почти исключительно фитопланктоном, кишечный тракт в 13 раз превышает длину тела рыбы.
Приемы добычи пищи разнообразны. Многие хищники непосредственно преследуют свою добычу, догоняя ее в открытой воде. Таковы акулы, жерех, судак. Есть хищники, подкарауливающие добычу и хватающие ее накоротке. В случае же неудачного броска они не делают попыток гнаться за добычей на большое расстояние. Так охотятся, например, щуки, сомы. Выше уже было указано, что пила-рыба и пилонос используют при охоте свой мечевидный орган. Они с большой скоростью врезаются в стаи рыб и делают «мечом» несколько сильных ударов, которыми убивают или оглушают жертву. Насекомоядная рыба брызгун (T.oxotes jaculator) имеет специальное приспособление, посредством которого она выбрасывает сильную струю воды, сбивающую насекомых с прибрежной растительности.
Многие донные рыбы приспособлены к раскапыванию грунта и выбиранию из него кормовых объектов. Карп способен доставать пищу, проникая в толщу грунта на глубину до 15 см, лещ — только до 5 см, окунь же практически вовсе не берет корм, находящийся в грунте. Успешно копаются в грунте американский многозуб (Polyodon) и среднеазиатский лопатонос (Pseudoscaphirhynchus), использующие для этого свой рострум (обе рыбы из подкласса хрящекостные).
Крайне своеобразно приспособление для добывания пищи у электрического угря. Эта рыба, перед тем как схватить свою добычу, поражает ее электрическим разрядом, достигающим у крупных особей 300 в. Угорь может производить разряды произвольно и несколько раз подряд.
Интенсивность питания рыб в течение года и вообще жизненного цикла неодинакова. Подавляющее большинство видов в период нереста перестают кормиться, сильно худеют. Так, у атлантического лосося масса мышц уменьшается более чем на 30 %. В связи с этим потребность в пище у них исключительно велика. Посленерестовый период носит название периода восстановительного питания, или «жора».
Размножение
Подавляющее большинство рыб раздельнополы. Исключение составляют немногие костные рыбы: морские окуни (Serranus scriba), дорады (Chrysophrys) и некоторые другие. Как правило, в случае гермафродитизма половые железы попеременно функционируют то как семенники, то как яичники, и самооплодотворение в силу этого невозможно. Только у морского окуня разные части гонады одновременно выделяют яйца и сперматозоиды. Иногда встречаются гермафродитные особи у трески, макрели, сельдей.
У некоторых рыб иногда наблюдается партеногенетическое развитие, которое, однако, не приводит к формированию нормальной личинки. У лососей неоплодотворенные икринки, заложенные в гнезде, не погибают и своеобразно развиваются до того времени, когда из оплодотворенных икринок выводятся эмбрионы. Это весьма своеобразное приспособление к сохранению кладки, так как если бы неоплодотворенные икринки ее развивались, а гибли и разлагались, то это привело бы к гибели всего гнезда (Никольский и Соин, 1954). У салаки и тихоокеанской сельди партеногенетическое развитие доходит иногда до стадии свободно плавающей личинки. Есть и другие примеры подобного рода. Однако ни в одном случае партеногенетическое развитие не приводит к формированию жизнеспособных особей.
У рыб известен и другой тип отклонения от нормального размножения, именуемый гиногенезом. В этом случае спермин проникают в яйцо, но слияния ядер яйцеклетки и спермия не происходит. У некоторых видов рыб развитие идет нормально, но в потомстве получаются только одни самки. Так бывает у серебряного карася. В Восточной Азии обнаруживаются как самки, так и самцы этого вида, и размножение протекает нормально. В Средней Азии, Западной Сибири и Европе самцы встречаются крайне редко, а в некоторых популяциях их нет вовсе. В таких случаях осеменение, приводящее к гиногенезу, осуществляется самцами других видов рыб (Н «Кольский, 1961).
Сравнительно с другими позвоночными, рыбы характеризуются громадной плодовитостью. Достаточно указать, что большинство видов откладывают в году сотни тысяч икринок, некоторые, например треска, до 10 млн., а луна-рыба даже сотни миллионов икринок. В связи со сказанным размер гонад у рыб вообще относительно велик, а ко времени размножения гонады еще резче увеличиваются. Нередки случаи, когда масса гонад в это время равна 25 и даже более процентов от общей массы тела. Громадная плодовитость рыб понятна, если учесть, что яйца у подавляющего большинства видов оплодотворяются вне тела матери, когда вероятность оплодотворения резко снижается. К тому же спермин сохраняют в воде способность к оплодотворению очень недолго: в течение короткого времени, хотя и различного в зависимости от условий, в которых происходит нерест. Так, у кеты и горбуши, нерестующей на быстром течении, где контакт спермы с икрой может осуществляться в очень малый промежуток времени, спермин сохраняют подвижность лишь на протяжении 10—15 секунд. У русского осетра и севрюги, нерестующих на более медленном течении, — 230 — 290 секунд. У волжской сельди через минуту после помещения спермы в воду подвижность сохраняло только 10% спермиев, а через 10 минут двигались лишь единичные сперматозоиды. У видов, нерестующих в относительно малоподвижной воде, спермин сохраняют подвижность дольше. Так, у океанической сельди спермин сохраняют способность к оплодотворению более суток.
Икринки, попадая в воду, продуцируют стекловидную оболочку, которая вскоре не дает возможности спермиям проникнуть внутрь. Все это уменьшает вероятность оплодотворения. Опытные подсчеты показали, что у лососей Дальнего Востока процент оплодотворенной икры равен 80%. У некоторых рыб этот процент еще меньше.
Кроме того, яйца развиваются, как правило, непосредственно в водной среде, они ничем не защищены и не охраняются. В силу этого вероятность гибели развивающейся икры, личинок и мальков рыб очень велика. Для промысловых рыб Северного Каспия установлено, что из всех выведшихся из икры личинок не больше 10% скатывается в море в виде сформировавшихся рыбок, остальные же 90% погибают (Никольский, 1944).
Процент рыб, доживающих до половозрелости, очень мал. Например, для севрюги он определяется в 0,01%, для осенней кеты Амура — 0,13— 0,58, для атлантического лосося — 0,125, для леща — 0,006—0,022% (Чефрас, 1956).
Таким образом, очевидно, что громадная начальная плодовитость рыб служит важным биологическим приспособлением для сохранения видов. Справедливость этого положения доказывается еще и ясной зависимостью между плодовитостью и условиями, при которых происходит размножение.
Наибольшей плодовитостью отличаются морские пелагические рыбы и рыбы, имеющие плавающую икру (миллионы икринок). Вероятность гибели последней особенно велика, так как она легко может быть съедена другими рыбами, выброшена на берег и т. п. Рыбы, откладывающие тяжелую, оседающую на дно икру, которая к тому же обычно приклеивается к водорослям или камням, имеют меньшую плодовитость. Многие лососевые откладывают икру в специально сооружаемые рыбами ямы, а некоторые засыпают потом эти ямы мелкой галькой. В этих случаях, следовательно, имеются первые признаки «заботы о потомстве». Соответственно с этим уменьшается и плодовитость. Так, семга выметывает от 6 до 20 тыс. икринок, кета — 2—5 тыс., а горбуша — 1—2 тыс. Укажем для сравнения, что севрюга откладывает до 400 тыс. икринок, осетры — 400—2500 тыс., белуга — 300—8000 тыс., судак — 300—900 тыс., сазан 400—1500 тыс., треска — 2500—10 000 тыс.
Трехиглая колюшка мечет икру в специальное гнездо, сооруженное из растений, и самец охраняет икру. Число икринок у этой рыбы равно 20— 100. Наконец, большинство хрящевых рыб, имеющих внутреннее осеменение, сложно устроенную оболочку яиц (которые они укрепляют на камнях или водорослях), откладывают яйца единицами или десятками.
У большинства рыб плодовитость с возрастом повышается и лишь к старости несколько снижается. Следует иметь в виду, что большинство наших промысловых рыб не доживают до возраста старения, так как к этому времени они бывают уже выловленными.
Как уже отчасти указано, для огромного большинства рыб характерно наружное оплодотворение. Исключение составляют почти все современные хрящевые рыбы и некоторые костистые. У первых в качестве копулятивного органа функционируют крайние внутренние лучи брюшных плавников, которые они при спаривании складывают вместе и вводят в клоаку самки. Много видов с внутренним оплодотворением среди отряда зубатых карпов (Cyprinodontiformes). Копулятивным органом у этих рыб служат видоизмененные лучи анального плавника. Внутреннее оплодотворение характерно для морского окуня (Sebastes marinus). Однако копулятивных органов у него нет.
В отличие от большинства позвоночных рыбы (если говорить о надклассе вообще) не имеют определенного сезона размножения. По времени нереста можно выделить по крайней мере три группы рыб:
1. Нерестующие весной и ранним летом — осетровые, карповые, сомовые, сельди, щука, окунь и др.
2. Нерестующие осенью и зимой — к ним принадлежат главным образом рыбы северного происхождения. Так, атлантический лосось начинает у нас нереститься с начала сентября; период икрометания растягивается у него в зависимости от возраста рыбы и условий водоема до конца ноября. Поздней осенью нерестится речная форель. Сиги мечут икру в сентябре — ноябре. Из морских рыб треска нерестится в финских водах с декабря по июнь, а у берегов Мурманска — с января до конца июня.
Как указывалось выше, у проходных рыб имеются биологические расы, отличные по времени захода в реки для нереста. Такие расы бывают, например, у кеты и семги.
3. Существует, наконец, третья группа рыб, которые не имеют определенного срока размножения. К ним принадлежат главным образом тропические виды, температурные условия обитания которых в течение года существенно не меняются. Таковы, например, виды семейства Cichlidae.
Места нереста крайне разнообразны. В море рыбы откладывают икру, начиная от зоны приливов и отливов, например пинагоры (Cyclopterus), атерина (Laurestes) и ряд других, и до глубин 500—1000 м, где нерестятся угри, некоторые камбалы и др.
Треска и морские сельди мечут икру у берегов, на сравнительно мелких местах (банках), но уже вне зоны приливов и отливов. Не менее разнообразны условия нереста в реках. Лещ в низовых ильменях Волги откладывает икру на водных растениях. Жерех, наоборот, выбирает места с каменистым дном и быстрым течением. В заводях, заросших водорослями, нерестятся окуни, которые прикрепляют икру к подводной растительности. На очень мелких местах, заходя в небольшие речки и канавы, нерестятся щуки.
Условия, в которых находится икра после оплодотворения, весьма разнообразны. Большинство видов рыб оставляют ее на произвол судьбы. Некоторые же помещают икру в специальные сооружения и более или менее длительное время ее охраняют. Бывают, наконец, случаи, когда рыбы носят оплодотворенную икру на своем теле или даже внутри своего организма.
Приведем примеры такой «заботы о потомстве». Нерестилища кеты расположены в мелких притоках Амура, в местах с галечным грунтом и относительно спокойным течением, глубиной 0,5—1,2 м; при этом важно наличие подземных ключей, дающих чистую воду. Самка, сопровождаемая одним или несколькими самцами, найдя место, пригодное для откладывания икры, ложится на дно и судорожно изгибаясь, расчищает его от травы и ила, поднимая при этом облако мути. Далее самка вырывает в грунте яму, что делается также ударами хвоста и изгибанием всего тела. После сооружения ямы начинается самый процесс нереста. Самка, находясь в яме, выпускает икру, а самец, находящийся рядом с ней, выпускает молоки. Около ямы обычно стоят несколько самцов, между которыми часто бывают драки.
Икра откладывается в яме гнездами, которых обычно бывает три. Каждое гнездо засыпается галькой, а когда закончено сооружение последнего гнезда, самка насыпает над ямой бугор овальной формы (длиной 2—3 м и шириной 1,5 м), который сторожит несколько дней, мешая другим самкам вырыть здесь яму для икрометания. Вслед за этим самка погибает.
Еще более сложное гнездо устраивает трехиглая колюшка. Самец вырывает на дне ямку, выстилает ее обрывками водорослей, затем устраивает боковые стенки и свод, склеивая растительные остатки клейким выделением кожных желез. В готовом виде гнездо имеет форму шара с двумя отверстиями. Затем самец загоняет в гнездо одну за другой самок и поливает каждую порцию икры молоками, после чего в течение 10—15 дней охраняет гнездо от врагов. При этом самец располагается относительно гнезда таким образом, что движения его грудных плавников возбуждают ток воды, идущий над икрой. Этим, видимо, обеспечивается лучшая аэрация, а следовательно, и более успешное развитие икры.
Дальнейшие усложнения описываемого явления «заботы о потомстве» можно видеть у рыб, которые носят оплодотворенную икру на своем теле.
У самки сомика аспредо (Aspredo laevis) кожа на брюхе в период нереста заметно утолщается и размягчается. После выметывания икры и оплодотворения ее самцом самка тяжестью своего тела вдавливает икру в кожу брюха. Теперь кожа имеет вид мелких сотов, в ячейках которых сидят икринки. Последние связаны с телом матери развивающимися стебельками, снабженными кровеносными сосудами.
У самцов иглы-рыбы (Syngnathus acus) и морского конька (Hippocampus) на нижней стороне тела имеются кожистые складки, образующие своего рода яйцевой мешок, в который самки откладывают икру. У морской иглы складки лишь загибаются на брюхо и прикрывают икру. У морского конька приспособление к вынашиванию развито еще сильнее. Края яйцевого мешка у него плотно срастаются, на внутренней поверхности образовавшейся камеры развивается густая сеть кровеносных сосудов, через посредство которых, видимо, и осуществляется газообмен эмбрионов.
Есть виды, вынашивающие икру во рту. Так бывает у американского морского сома (Galeichthys fells), у которого самец вынашивает в ротовой полости до 50 яиц. В это время он, видимо, не питается. У других видов (например, рода Tilapia) икру носит во рту самка. Иногда во рту бывает более 100 икринок, которые приводятся самкой в движение, что связано, видимо, с обеспечением лучшей аэрации. Инкубационный период (судя по наблюдению в аквариуме) длится 10—15 дней. В это время самки почти не питаются. Любопытно, что и после вывода мальки еще некоторое время при опасности прячутся в рот матери.
Упомянем о весьма своеобразном размножении горчака (Rhodeus sericeus) из семейства карповых, широко распространенного в России. В период нереста у самки развивается длинный яйцеклад, которым она откладывает икру в мантийную полость моллюсков (Unio или Anodonta). Здесь яйца оплодотворяются сперматозоидами, всасываемыми моллюсками с током воды через сифон. (Самец выделяет молоки, находясь рядом с моллюском.) Эмбрионы развиваются в жабрах моллюска и выходят в воду, достигнув длины около 10 мм.
Последняя степень усложнения процесса размножения у рыб выражается в живородности. Оплодотворенная в яйцеводах, а иногда даже в яичниковом мешке икра не поступает в наружную среду, а развивается в половых путях матери. Обычно развитие осуществляется за счет желтка яйца, и лишь в конечных стадиях эмбрион питается и за счет выделения стенками яйцевода особой питательной жидкости, воспринимаемой эмбрионом через рот или через брызгальце. Таким образом, описываемое явление правильнее обозначать как яйцеживорождение. Однако у некоторых акул (Charcharius Mustelus) формируется своеобразная желточная плацента. Она возникает путем установления тесной связи между богатыми кровеносными сосудами выростами желточного пузыря и такими же образованиями стенок матки. Через эту систему осуществляется обмен веществ у развивающегося эмбриона.
Яйцеживорождение в наибольшей мере характерно для хрящевых рыб, у которых оно наблюдается даже чаще, чем откладывание яиц. Наоборот, среди костных рыб это явление наблюдается очень редко. В качестве примера можно указать на байкальских голомянок (Comephoridae), морских собачек (Blenniidae), морских окуней (Serranidae) и особенно зубатых карпов (Cyprinodontidae). Все яйцеживородящие рыбы имеют малую плодовитость. Большинство рождают единицы детенышей, реже десятки. Исключения очень редки. Так, например, морская собачка рождает до 300 молодых, а норвежская морулъка (Blenniidae) даже до 1000.
Мы привели ряд случаев, когда оплодотворенная икра не оставляется на произвол судьбы и рыбы проявляют в том или ином виде заботу о ней и о развивающейся молоди. Такая забота свойственна ничтожному меньшинству видов. Основным, в наибольшей мере характерным типом размножения рыб является такой, при котором икра оплодотворяется вне тела матери и в последующем родители оставляют ее на произвол судьбы. Именно этим и объясняется огромная плодовитость рыб, обеспечивающая сохранение видов и при очень большой, неизбежной в указанных условиях гибели икры и молоди.
Рост и возраст
Продолжительность жизни рыб весьма различна. Есть виды, живущие немногим более года: некоторые бычки (Gobiidae) и светящиеся анчоусы (Scopelidae). С другой стороны, белуга доживает до 100 и более лет. Однако в связи с интенсивным промыслом реальная продолжи-тельность жизни измеряется немногими десятками лет. Некоторые камбалы живут 50—60 лет. Во всех этих случаях имеется в виду предельная потенциальная продолжительность жизни. В условиях же регулярного промысла действительная продолжительность жизни много меньше.
В отличие от большинства позвоночных, как правило, рост рыбы не прекращается по достижении половой зрелости, а продолжается в течение большей части жизни, до периода старости. Наряду со сказанным для рыб характерна четко выраженная сезонная периодичность роста. Летом, особенно в период нагула, они растут значительно быстрее, чем в малокормный зимний период. Эта неравномерность роста сказывается на строении ряда костей и чешуи. Периоды замедленного роста отпечатываются на скелете в
виде узких полос или колец, состоящих из мелких клеток. При рассмотрении в падающем свете они кажутся светлыми, в проходящем свете, наоборот, темными. В периоды усиленного роста откладываются широкие кольца или слои, которые в проходящем свете кажутся светлыми. Комбинация двух колец — узкого зимнего и широкого летнего — и представляет годовую отметку. Подсчет этих отметок позволяет определить возраст рыбы.
Определение возраста производится по чешуе и некоторым частям скелета.
Так, по чешуе можно установить число прожитых лет у лососевых, сельдевых, карповых, трески. Чешуя промывается в слабом растворе нашатырного спирта и просматривается между двумя предметными стеклами под микроскопом и лупой. У окуневых, налима и некоторых других рыб возраст устанавливается по плоским костям, например по жаберной крышке и клейтруму. У камбал и тресковых рыб для этой цели служат отолиты, которые предварительно обезжириваются, а иногда и шлифуются.
Возраст осетровых, сома и некоторых акул устанавливается рассмотрением поперечного среза луча плавника: у акул — непарного, у осетровых — грудного.
Определение возраста рыб имеет огромное теоретическое и практическое значение. При рационально поставленном промысле анализ возрастного состава улова служит важнейшим критерием для установления перелова или недолова. Увеличение плотности тела младших возрастов и уменьшение старших указывает на напряженность промысла и угрозу перелова. Наоборот, большой процент рыб старших возрастов говорит о неполном использовании запасов рыбы. «Так, например, если в улове воблы (Rutilus rutilus caspius) большое количество семи- и восьмилетних особей будет указывать, как правило, на недолов (вобла обычно становится половозрелой по достижении трехлетнего возраста), то наличие в улове осетра (Acipenser gtildenstadti) особей главным образом в возрасте 7—8 лет будет указывать на катастрофическое положение промысла (осетр становится половозрелым не раньше 8—10-летнего возраста), так как в исследованном улове осетра преобладают неполовозрелые особи» (Никольский, 1944). Кроме того, сопоставляя возраст и размеры рыб, можно сделать важные заключения о темпах их роста, часто связанных с кормностью водоемов.