
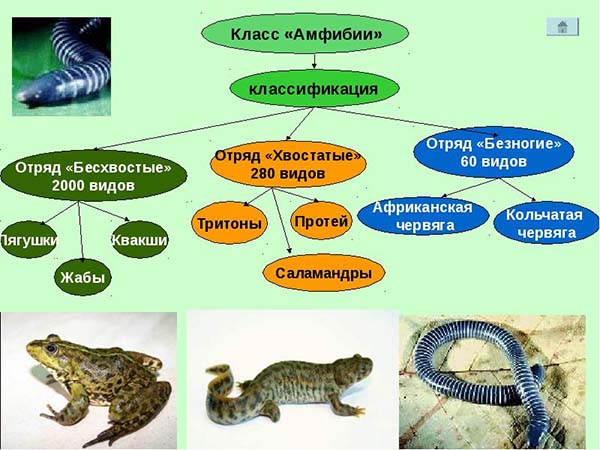
Систематика современных земноводных
Отряд 1 Хвостатые амфибии (Caudata, или Urodela)
Характеризуются удлиненным, вальковатым телом с соразмерно развитыми головным, туловищным и хвостовым его отделами. Хвост в поперечном сечении круглый или сжатый с боков, иногда с кожной оторочкой в виде киля. Конечности у большинства хорошо развиты, при этом передняя и задняя пары обыкновенно одинаковой длины. У некоторых видов нет задних конечностей (сирены).
Позвонки амфицельные или опистоцельные. К туловищным позвонкам причленяются зачаточные верхние ребра. Костный череп более дифференцирован, чем у бесхвостых. Лобные и теменные кости не слиты, имеются парные глазоклиновидные кости. Ключицы отсутствуют. В отличие от бесхвостых, предплечье и голень имеют типичное строение и состоят из двух костей (лучевой и локтевой, большой и малой берцовой).
Кровеносная система устроена сравнительно просто. Более или менее полная перегородка в предсердии свойственна только высшим хвостатым. Сохраняются все четыре дуги аорты. Наряду с задней полой веной сохраняются и кардинальные вены. У некоторых пожизненно сохраняются жабры, чаще наружные, реже внутренние. Кожное дыхание развито очень сильно.
Барабанной полости и барабанной перепонки нет. У многих хвостатых, в отличие от других отрядов амфибий, пожизненно сохраняются органы боковой линии.
Оплодотворение у многих видов внутреннее. Размножение происходит путем откладывания оплодотворенной икры, личинок или живорождением. У ряда видов наблюдается способность размножаться в личиночной стадии (неотения).
Общее число современных видов равно примерно 280. Распространены большей частью в умеренном поясе западного и восточного полушарий. В южном полушарии практически отсутствуют.
Наибольшее число видов относится к семейству саламандровых (Salamandridae). Форма тела у них ящерецевидная, хвост длинный, округлый или сжатый с боков. Две пары конечностей развиты сравнительно хорошо. Личинки дышат наружными жабрами, взрослые жабр не имеют и дышат легкими и через кожу.
Обычными видами этого семейства являются тритоны, распространенные в Европе, на Кавказе, в Сибири и местами в Средней Азии. Обыкновенный и гребенчатый тритоны (Triturus vulgaris) и (Triturus cristatus) населяют летом небольшие стоячие или медленно текущие водоемы с более или менее развитой растительностью. В водоемах происходит размножение и развитие личинок. В конце лета тритоны покидают водоемы и держатся под стволами деревьев, под камнями или в трещинах земли. Зимуют на суше под пнями, в дуплах лежащих деревьев, в кучах листьев. Весной, когда местами еще лежит снег, тритоны вновь переселяются в воду. Питаются животной пищей.
Несколько видов настоящих саламандр (Salamandra) распространено в Средней и Южной Европе, на Западном Кавказе, в Малой Азии и Северной Африке. Саламандры характеризуются неуклюжим сложением тела, крупным без киля хвостом и обычно пятнистой окраской.
Пятнистая, или огненная, саламандра (S. salamandra) обитает в сырых темных лесах Европы, Северной Африки и Малой Азии. Днем держится под корнями деревьев, под камнями или в норах. Активна обычно ночью и только после дождей выходит из убежищ днем. С водой связана меньше, чем тритоны. Оплодотворение чаще бывает на суше; при этом самец и самка сближают клоаки и сперматофоры переходят в половые пути самки. Бывают случаи оплодотворения и в воде. Пятнистая саламандра обычно рождает личинок с жабрами, которые заканчивают метаморфоз в водоемах.
В Северной Америке широко распространены виды семейства амбистом (Ambystoma). Это сравнительно стройные животные с гладкой кожей, которая имеет вертикальные борозды, отчего туловище кажется кольчатым. Хвост толстый, без кожистой оторочки. Общая окраска буро-коричневая с мелкими светлыми пятнами. Длина тела 15—23 см. Личинки амбистом, известные под названием аксолотлей, способны размножаться (неотения).
Своеобразные безлегочные саламандры (Plethodontidae) распространены преимущественно в Северной Америке, но некоторые виды живут в Центральной и Южной Америке, а один или два вида — в Южной Европе. Все виды утратили легкие, что было связано с переходом к обитанию только в воде. Однако многие виды, особенно в тропиках и пещерах, вновь стали наземными. У всех видов дыхание осуществляется через кожу и отчасти через слизистую глотки. Наземные виды яйца откладывают часто вне водоема: в норах или под корнями, а иногда в дуплах. Самка обвивается вокруг комка яиц и не покидает его до выхода из них молодых.
К семейству амфиумовых (Amphiumidae) принадлежат одни из наиболее крупных современных амфибий. Длина тела некоторых видов достигает одного метра. Конечности у них развиты слабо и почти непригодны для хождения. Органами дыхания наряду с легкими служат внутренние жабры, в которые ведут жаберные щели (по одной с каждой стороны).
На юго-востоке Северной Америки распространена амфиума (Amphiuma means). Тело ее угревидной формы, длиной 70—100 см. Конечности рудиментарны и совершенно не годны для передвижения по суше. Большую часть жизни амфиумы проводят в воде — в болотах, канавах и медленно текущих ручьях. Яйца откладывают на суше в сырых местах, у воды. Самка свертывается спирально вокруг комка яиц и держится так почти весь период развития яиц.
В семействе протеевых (Proteidae) лишь два современных вида. Один из них распространен в подземных водах Балканского полуострова, другой — в мелких заросших водоемах Северной Америки. Протеи в течение всей жизни сохраняют наружные пучкообразные жабры. Глаза скрыты под кожей. Конечности очень маленькие, непригодные для ходьбы. Особи, населяющие подземные водоемы, имеют кожу, лишенную пигмента, и окрас-ка их мясо-красная, обитающие в открытых водоемах (в Америке) имеют серую или бурую с темными пятнами окраску. Длина тела 25—35 см. Размножаются путем откладывания оплодотворенных яиц. У балканского протея наблюдали случаи живорождения. Последнее, видимо, бывает при понижении температуры воды ниже 15° С.
Семейство сирен (Sirenidae) включает два вида распространенных в Северной Америке. Как и у протеев, у них всю жизнь сохраняются наружные жабры, кроме того, имеются рудиментарные жаберные щели. Задние конечности отсутствуют. Передние конечности развиты слабо. Глаза затянуты кожей. Общая форма тела угревидная, длиной 60—70 см. Населяют болота, где держатся под кочками и пнями.
Семейство углозубов (Hynobiidae) включает весьма примитивные виды (около 20), которые имеют амфицельные позвонки и наружное оплодотворение. Распространено это семейство преимущественно в таежной полосе Восточной Азии. Типичный вид — сибирский углозуб (Hynobius keyserlingi) распространен от Камчатки на запад до республики Коми и Новгородской области. В водоемах живет только в период размножения. Характерна большая устойчивость к низким температурам: даже при температуре около 2—4° С углозубы остаются активными.
К семейству скрытожаберных (Cryptobranchidae) принадлежат наиболее примитивные современные хвостатые. Оплодотворение у них наружное. Для примера укажем на японскую гигантскую саламандру (Меgalobatrachus japonicus), которая имеет длину тела около 150 см. Обитает она в горных реках острова Ниппон (Япония) и в реках Центрального Китая. Этот вид держится под камнями и навесами подмытых берегов. Питается рыбами, лягушками, червями. Яйца откладывает в воде, в норах на участках с тихим течением.
Мясо этой саламандры употребляют в пищу и за ней усиленно охотятся.
Отряд 2 Безногие амфибии (Apoda)
Весьма своеобразная группа крайне специализированных и в то же время примитивных амфибий. Специализация связана с подземным, роющим образом жизни. Общая форма тела червеобразная, у многих видов с кольцевидными перетяжками, что придает этим животным внешнее сходство с земляными червями. Ног нет. Кожа голая, очень богатая железами, и тело обильно покрыто слизью. Хвост очень короткий, и отверстие клоаки открывается почти у самого конца тела. В связи с роющим образом жизни глаза рудиментарные, слуховые нервы развиты слабо, барабанной перепонки нет.
Имеется копулятивный орган — выпячивание стенки клоаки.
Безногие обладают рядом примитивных особенностей строения. Так, в коже у них имеются островки костной ткани, которые считают остатками панциря стегоцефалов. Покровные кости черепа развиты значительно сильнее, чем у других земноводных, что также сближает безногих с панцирными амфибиями. Позвонки амфицельные. Есть настоящие ребра. Слуховая косточка — стремя — еще сочленяется с квадратной костью. Перегородка между предсердиями недоразвита.
Отряд содержит одно семейство — червяг (Coecieliidae), насчитывающее около 60 видов, распространенных главным образом в Южной Америке, отчасти в тропической Африке и на юге Азии. Кольчатая червяга (Siphonops annulatus), распространенная в Южной Америке, имеет длину около 40 см. Живет во влажных местах и в землю зарывается на глубину до 50 см.
В Индостане, Индокитае, на Цейлоне и на Больших Зондских островах распространен другой вид — цейлонский рыбозмей (Ichthyophis glutinosus). Длина его около 40 см. Держится в почве на глубине 20—30 см по берегам рек и озер.
Только один род — Typhleonect.es — населяет пресные водоемы Южной Америки.
Размножаются безногие чаще путем откладывания яиц (20—30 штук), которые помещаются в земляных норах, под пнями. Самки обычно обвиваются вокруг яиц и обильным выделением своих кожных желез предохраняют яйца от высыхания.
Выведшиеся личинки мигрируют в воду, где заканчивают развитие. Наружные жабры личинки теряют, еще находясь в яйце, и период их пребывания в воде короток. Некоторые виды живородящи. Оплодотворение у всех внутреннее. Питаются почвенными беспозвоночными. Некоторые виды живут в гнездах муравьев, личинками которых и питаются.
<
Отряд 3. Бесхвостые амфибии (Ecaudata, или Anura)
Бесхвостые — наиболее высокоорганизованная, многочисленная и широко распространенная группа современных амфибий. К ней принадлежит около 1300 видов, обитающих на всех материках, кроме Антарктического. Особенно многочисленны бесхвостые в Южной Америке. Несмотря на обилие видов, все бесхвостые довольно однообразны: форма тела их лягвообразная, без наружного хвоста; всегда имеются хорошо развитые передние и задние конечности, причем задние лапы развиты сильнее передних. Этим обусловливается характерное скачкообразное передвижение по земле.
В скелете характерно: продельная форма позвонков (у большинства видов), отсутствие ребер (за одним исключением), срастание лобных и теменных костей, а также лучевой и локтевой, большой и малой берцовой костей. Последнее связано с указанным выше типом движения по твердому субстрату. В этой же связи частично срастаются кости запястья и предплюсны. Барабанная полость развита. У взрослых никогда не сохраняются наружные жабры или жаберные щели. Осеменение обычно наружное, и копулятивных органов нет.
Многочисленные виды бесхвостых амфибий распределяются по значительному числу семейств, из которых ниже указаны лишь некоторые.
К наиболее древнему и примитивному семейству леопельмовых (Leopeknidae) относятся только два вида, один из которых распространен в Северной Америке, другой в Новой Зеландии. Самцы этих видов пожизненно сохраняют наружный хвост, при этом у новозеландского вида скелет хвостового отдела позвоночника сегментирован. Позвонки амфицельны. Большую часть времени они проводят в водоемах.
Виды семейства круглоязычных (Discoglossidae) характерны круглым без вырезки языком. В нашей фауне встречаются жерлянки (Bombina) — маленькие лягушки длиной 5 см. Верх тела у них темно-оливковый, почти черный, низ очень яркий — из смеси оранжевых и черных пятен. Железы кожи выделяют едкий секрет. Держатся в небольших водоемах, часто в лужах. Распространены в средней полосе Европейской части России и на юге Дальнего Востока.
К этому же семейству принадлежит и так называемая жаба-повитуха (Alytes) — небольшая лягушка (около 5 см) пепельно-серого цвета, распространенная в Западной Европе. Биологически интересна особенностями размножения. Самка откладывает около 50 яиц, которые соединены в виде четкообразного шнура длиной примерно 1 м. После оплодотворения икры самец наматывает ее на задние лапы и держится на суше в тенистых местах. Через 17—18 дней самец переходит в воду. Развившиеся к тому времени личинки покидают яйцевые оболочки и, как и головастики других лягушек, ведут водный образ жизни.
Жабы (сем. Bufonidae) сравнительно мало связаны с водой и распространены даже в пустынных областях. Это связано с тем, что большинство видов, будучи ночными животными, избегают дневной жары. Кроме того, клетки верхнего слоя эпидермиса кожи частично ороговевают. У некоторых жаб выработались приспособления к размножению вне водоемов. Так, австралийская жаба псевдофрина откладывает яйца (икру) на землю во влажных местах. Южноамериканские жабы циклорамфус и некоторые другие виды помещают яйца между камнями или зарывают их в почву. Головастиков у этих видов не бывает, и из яйца, богатого желтком, вылупляются вполне оформившиеся лягушки.
Два вида жаб, обыкновенная, и зеленая (Bufo bufo и В. viridis), распространены в нашей стране преимущественно в средних и южных ее широтах. Зеленая жаба есть и в равнинной Средней Азии, где обитает в оазисах. Оба вида откладывают яйца в воду. Полезны истреблением насекомых и моллюсков. (Два других вида жаб нашей фауны — монгольская и камышевая — имеют в России весьма ограниченное распространение.).
Многочисленные виды семейства квакши (Hylidae) — мелкие лягушки, ведущие в той или иной мере древесный образ жизни. На концах пальцев, а у некоторых и на брюхе имеются присоски, при помощи которых они удерживаются на стволах деревьев, на ветвях и других предметах. Распространены преимущественно в Южной Америке и Австралии. Два вида — обыкновенная и дальневосточная квакши (Hyla arborea и Н. japonica) — встречаются и у нас: первая — на Украине, в Крыму, на Кавказе, вторая — на Дальнем Востоке. Обитают в лесах, предпочтительно широколиственных, реже в тростниках или среди другой высокой травянистой растительности. Вне периода размножения квакша много времени проводит на деревьях, по которым лазает с удивительной быстротой и ловкостью. Икрометание происходит в воде.
Сумчатые квакши Южной Америки (Nototrema) характерны тем, что у самок этих лягушек на спине имеется кожистый карман, в который самец помещает оплодотворенные яйца. Развитие молоди происходит в сумке, причем у одних видов головастики уходят в воду и там заканчивают свое превращение, у других видов все стадии превращения происходят в сумке, и из нее выходят уже сформировавшиеся четвероногие лягушки. Южноамериканские квакши филломедузы (Phyllomedusa) размножаются на деревьях и яйца откладывают в свернутые трубочкой листья. Другие виды тропических квакш откладывают икру в воду, скопившуюся в дупле, в пазухах листьев, на пнях.
Настоящие лягушки (сем. Ranidae) распространены по всем материкам, исключая Австралию и Антарктиду. Большинство видов размножается в воде, и личинки претерпевают в водоемах полное превращение.
Ряд видов распространен в нашей стране. Таковы озерная лягушка (Rana ridibunda) и близкая к ней прудовая лягушка (R. esculenta). Распространены в низовьях Днепра, Дона, Волги, Урала. Они очень связаны с водой и никогда не отходят далеко от водоемов. Местами приносят незначительный вред, уничтожая мальков рыб.
Значительно менее с водой связаны травяная лягушка (R. temporaria), распространенная в Европе, Западной Сибири (на восток до Оби) и на Дальнем Востоке, и остромордая лягушка (R. terrestris), распространенная очень широко, на север местами до полярного круга, а на восток до бассейна Амура. В водоемах они держатся только во время размножения. Летом живут на суше, часто далеко от ближайшего водоема. Полезны истреблением вредных насекомых.
Хотя большинство настоящих лягушек — наземные или полуводные жители, есть виды, ведущие древесный образ жизни. Такова, например, яванская летающая лягушка (Rhacophorus reinwardti). Большую часть времени она проводит на деревьях, по которым хорошо лазает и легко пере¬прыгивает с дерева на дерево. Прыжок облегчается большими перепонками между пальцами, которые лягушка при прыжке широко раздвигает. При длине лягушки около 7 см поверхность перепонок между растопыренными пальцами равна примерно 20 см2. Планируя, лягушка может перелетать расстояние в 10—15 м.
Древесные лазающие, но не летающие лягушки есть также в Африке; такова, например, хватающая лягушка (Chiromantis), откладывающая икру на ветвях деревьев и на листьях, расположенных над водой. Вылупляющиеся личинки падают в воду и там заканчивают свое развитие.
В тропиках Южной Америки распространена очень интересная лягушка — суринамская пипа (Pipa americana), принадлежащая к особому эндемичному для Южной Америки семейству пиповых (Pipidae). Эта лягушка живет преимущественно в водоемах, и органы боковой линии у нее сохраняются и во взрослом состоянии. Крайне своеобразно размножение пипы.
Происхождение земноводных
Происхождение земноводных представляет исключительный интерес. В данном случае мы имеем дело не только с возникновением еще одного нового класса животных, но и с появлением позвоночных в новой, наземной среде обитания. Выход позвоночных из водной среды обитания на сушу привел их к столкновению с новыми и значительно более разнообразными условиями жизни, обусловил новые, сложные пути их эволюционного развития, приведшие к появлению в последующем высших групп: рептилий, птиц и млекопитающих.
Переход позвоночных от водного к наземному образу жизни должен был сопровождаться появлением в основном двух решающих приспособлений: дыхания кислородом атмосферы и передвижения по твердому субстрату. Другими словами, жаберное дыхание должно было замениться легочным, а конечности типа плавника — пятипалыми конечностями, представляющими многочленный рычаг, служащий для опоры тела о твердый субстрат. Совершенно очевидно, что параллельно должны были измениться и другие системы органов: кровообращения, органов чувств, нервной системы и др.
Намеки на появление указанных приспособлений мы встречаем среди разнообразных групп рыб. Известны случаи, когда рыбы на то или иное время выходят из воды и кровь у них частично окисляется за счет кислорода атмосферы. Таков, например, ползун (Anabas), который, выходя из воды, даже взбирается на деревья и сидит там подолгу. Выползают на сушу не¬которые бычки (Gobiidae) и прыгуны (Periophthalmus). Последние свою добычу ловят чаще на суше, чем в воде. Хорошо известна способность находиться вне воды некоторых двоякодышащих. Однако все эти приспособления количественно столь незначительны, что не позволяют появиться принципиально новому биологическому качеству. Предков земноводных надо искать среди других, менее специализированных групп пресноводных рыб.
Как указывает акад, И. И. Шмальгаузен (1964), в конце девона в пресных водоемах появились первые амфибии — ихтиостегиды (Ichthyostegidae), они были настоящими переходными формами между кистеперыми рыбами и земноводными. Так, у них имелись рудименты жаберной крышки, настоящий рыбий хвост. Кожа была покрыта мелкой рыбьей чешуей. Однако наряду с этим они имели парные конечности типа пятипалых конечностей наземных позвоночных. Ихтиостегиды жили не только на суше, но и в воде. Можно предположить, что они не только размножались, но и кормились в воде, систематически выползая на сушу.
В дальнейшем в каменноугольном периоде возник ряд ветвей, которым придают таксономическое значение надотрядов или отрядов. Надотряд лабиринтодонтов (Labyrinthodontia) был наиболее разнообразен. Ранние формы имели сравнительно небольшие размеры и рыбообразную форму тела. Более поздние достигали весьма крупных размеров (длина черепа 1 м и более), тело их было уплощено и заканчивалось коротким толстым хвостом. Лабиринтодонты существовали до конца триаса.
Надотряд прыгающих (Salientia) включает и всех современных бесхвостых (отряд Anura). Ископаемый предок бесхвостых выделен в особый отряд (Pronura). Он известен примерно с перми, но бесхвостые более или менее современного тина появились, видимо, только в меловое время.
В карбоне возникла и вторая основная ветвь первичных амфибий — лепоспондилы (Lepospondyli). Они имели мелкие размеры и были хорошо приспособлены к жизни в воде. Некоторые из них вторично утратили конечности. Начальные их формы просуществовали до середины пермского периода. Полагают, что они дали начало отрядам современных амфибий — хвостатых (Caudata) и безногих (Apoda).
Все эти ранние вымершие амфибии, которых иногда именуют стегоцефалами (панцирноголовыми), характеризовались сплошным панцирем из кожных костей, покрывавшим черепную коробку сверху и с боков, так что оставались отверстия только для ноздрей, глаз и теменного органа. Кроме того, у многих был брюшной панцирь из налегающих друг на друга костных чешуй. Предками стегоцефалов, несомненно, были костные рыбы, сочетавшие примитивные особенности организации (например, слабое окостенение первичного скелета) с наличием дополнительных к жабрам органов дыхания в виде легочных мешков и такими парными конечностями, которые могли служить не только для гребли, но и для опоры о твердый субстрат и из которых, следовательно, могли бы развиться пятипалые конечности. Лучеперые рыбы (Actinopterygii) не обладают этими признаками. Двоякодышащие (Dipnoi), хотя и имеют легочное дыхание, представляют весьма специализированную группу донных рыб. Достаточно указать на такие признаки, как бисериальные плавники, отсутствие вторичных верхних челюстей, наличие роговых зубов и т. д. Наиболее близко к стегоцефалам стоят кистеперые рыбы (Crossopterygii). Они обладали легочным дыханием, их конечности имели скелет, в основном сходный с таковым стегоцефалов. Проксимальный отдел состоял из одной кости, соответствующей плечу или бедру, следующий сегмент состоял из двух костей, соответствующих предплечью или голени; далее располагался отдел, состоявший из нескольких рядов костей, он соответствует кисти или стопе. Обращает, наконец, внимание удивительное сходство в расположении покровных костей черепа у древних кистеперых и стегоцефалов. Надо при этом иметь в виду, что известные науке кистеперые рыбы не могли быть предками наземных позвоночных, так как жили они в более позднее время или были слишком специализированы. Начало наземным позвоночным дали более древние кистеперые, еще не обнаруженные палеонтологией.
Выход позвоночных на сушу явился результатом длительного приспособления кистеперых рыб к жизни в водоемах, бедных кислородом. Девонский период, в котором возникли стегоцефалы, видимо, характеризовался сезонными засухами, во время которых жизнь во многих пресных водоемах была для рыб затруднительна. Обеднению воды кислородом и затрудненности плавания в ней способствовала обильная растительность, произраставшая в каменноугольное время по болотам и берегам водоемов. Растения падали в воду, и в этих условиях могли возникнуть приспособления рыб к дополнительному дыханию легочными мешками. Само по себе обеднение воды кислородом еще не являлось предпосылкой для выхода на сушу. В этих условиях кистеперые рыбы могли подниматься на поверхность и заглатывать воздух. Но при сильном усыхании водоемов жизнь для рыб становилась уже невозможной. Неспособные к передвижению по суше, они погибали. Только те из водных позвоночных, которые одновременно со способно¬стью к легочному дыханию приобрели ко¬нечности, могущие обеспечить передвиже¬ние по суше, могли переживать эти усло¬вия. Они выползали на сушу и переходили в соседние водоемы, где еще сохранялась вода. Вместе с тем передвижение по суше для животных, покрытых толстым слоем костной чешуи, было затруднено и костный панцирь на теле не обеспечивал возможности кожного дыхания, столь характерного для всех амфибий. Указанные обстоятельства, видимо, явились предпосылкой редукции костного панциря на большей части тела. У стегоцефалов (и то не у всех) он сохранился (не считая панциря черепа) только на брюхе. Двигаясь по суше, они волочили тело по земле, и брюшной панцирь служил надежной защитой тела от механических повреждений.
Стегоцефалы дожили до начала мезозоя. Современные же отряды амфибий появляются только в конце мезозоя (конец юры — начало мела). Таким образом, непосредственной связи между ними установить пока не удается. В общих чертах можно указать, что исходной группой для хвостатых и бесхвостых был отряд Phyllospondyli. Ближе к ним стоят бранхиозавры. Однако разделение хвостатых и бесхвостых относится, видимо, к концу палеозоя. О предках безногих ничего достоверного неизвестно.
Экология амфибий
Условия существования и общее распространение,. Жизненная обстановка амфибий весьма разнообразна. Среди них есть чисто водные формы, никогда не выходящие на сушу. Большинство их принадлежит к хвостатым амфибиям; таковы протеи, сирены, некоторые амфиумовые. Только немногие бесхвостые также живут исключительно в воде. Таковы, например, африканские шпорцевые лягушки.
Большинство бесхвостых ведут полуводный образ жизни. В период размножения они живут в водоемах. Многие зимуют также в воде. Вне указанного времени эти виды живут на суше и часто уходят от водоемов на большие расстояния. Таковы жабы, бурые лягушки, огненная и кавказская саламандры. Зеленые лягушки также много времени проводят вне воды, но от водоемов далеко не уходят и при опасности одним прыжком скрываются в воду.
Среди бесхвостых есть виды, живущие главным образом на деревьях. В большинстве это обитатели тропических лесов, которые и размножаются на деревьях, используя для откладывания яиц воду, скопившуюся в дуплах и на крупных листьях. Степень связанности с деревьями и приспособленности к древесному образу жизни у них различна. Так, наша квакша много времени проводит на деревьях, но размножается на земле, в водоемах. Квакши лазают по деревьям при помощи больших округлых присосок, расположенных на концах пальцев и богатых железами, выделяющими клейкий секрет. У других видов клейкий секрет свойствен и некоторым участкам брюшной поверхности. У филломедуз пальцы приспособлены для обхватывания ветвей. Есть сведения, что, если животное это ухватилось за ветку, его невозможно оторвать, не повредив лапку. У африканских хватающих лягушек (Chiromantis) специализация лап еще большая: два внутренних пальца у них могут противополагаться остальным, образуя своего рода клешни. Упомянем, наконец, яванскую летающую лягушку (Rhacophorus), у которой между удлиненными пальцами расположены перепонки, служащие для планирования.
Последняя основная биологическая группа амфибий — это роющиеся в земле виды. К ним относятся почти все безногие, представляющие в значительной мере обитателей толщи почвы, редко выходящих днем на поверхность. Среди бесхвостых также есть виды, могущие глубоко закапываться в землю. Такова, например, чесночница. Однако для подавляющего большинства видов лягушек почва служит лишь местом временного нахождения, а не постоянной средой обитания, как для безногих наземных земноводных.
Хотя амфибии, как это показано выше, живут в разнообразных сре¬дах, их общее географическое распространение связано с наличием весьма специфических жизненных условий. Основные из них следующие: тепло, наличие водоемов, значительная влажность воздуха, химизм воды и почвы.
Тепло — обязательное условие успешного существования амфибий. При температуре +7, +8° С большинство видов уже впадают в оцепенение, а при температуре —2° С они погибают. Низкая температура воды не обеспечивает развития яиц и личинок. В указанной связи амфибии наиболее многочисленны в тропических областях. По мере движения к полюсам их становится все меньше. За Северный полярный круг незначительно переходят (и то в немногих местах) только травяная и остромордая лягушки и сибирский четырехпалый тритон.
Засушливый жаркий климат весьма неблагоприятен для амфибий. Имея тонкую кожу, они теряют много влаги из тела. П. В. Терентьев (1959) указывает, что если лягушки высыхают быстро, то они умирают при потере 15% первоначальной массы; но при медленном высушивании выдерживают почти вдвое большую потерю массы (по данным А. Г. Банникова, квакши — до 75%). Верхний температурный предел жизнедеятельности для большинства видов лягушек равен примерно +40° С.
Лучше других засушливые условия переносят жабы, что связано с появлением у них ороговения эпидермиса, защищающего тело от иссушения. Кроме того, жабы ведут ночной образ жизни и в силу этого избегают иссушающего действия дневного зноя. В связи со сказанным жабы обитают и в пустынных областях Средней Азии, где они встречаются, однако только в оазисах.
У австралийской циклораны в мочевом пузыре содержится запас воды, который составляет до 50% массы тела.
Сухость воздуха имеет значение не только для эффективности дыхания, но и для температуры тела.
Амфибии не могут жить в соленой воде или на сильно засоленных почвах. Установлено, что раствор хлористого натра концентрацией менее 1% губителен для личинок земноводных и для многих взрослых форм. В силу сказанного морские проливы представляют непреодолимую преграду для расселения земноводных, и по этой причине они отсутствуют на большинстве океанических островов, даже в тропических странах с их оптимальными для существования этих животных условиями.
Защитные приспособления у амфибий развиты сравнительно слабо, и в подавляющем большинстве случаев действие их не имеет активного характера.
Наиболее действенным защитным приспособлением, видимо, являются кожные железы, секрет которых ядовит. Попадая на слизистые оболочки, он вызывает сильное раздражение, а иногда и отравление. Наблюдались случаи, когда лягушки (Rana), посаженные в одну банку с жерлянками (Bombina), уже через несколько часов погибали, будучи отравленными секрецией кожных желез жерлянок. Еще сильнее действует этот секрет, если его ввести в кровяное русло. При значительной дозе собака, например, погибает менее чем через час. Особенно сильно развиты ядовитые железы у жаб, жерлянок, чесночниц и саламандр. Видимо, по этой причине их почти не трогают птицы и звери.
Известное значение имеет покровительственная окраска. У одних видов — саламандр, жерлянок — большая или меньшая часть тела окрашена очень ярко и, видимо, имеет «предостерегающее» значение. Зеленые и бурые лягушки окрашены так, что они малозаметны в окружающей обстановке. Некоторые виды, например квакша, меняют окраску в зависимости от условий места нахождения. Так, на листьях она ярко-зеленая, на стволах деревьев — бурая.
Активно защищаются лишь немногие виды. Крупные лягушки, например рогатые, бросаются на преследователя и делают попытку кусаться.
Упомянем, наконец, о способности к регенерации. Некоторые саламандры, будучи схвачены за хвост, отламывают его. В последующем хвост отрастает вновь — регенерирует. Установлено, что регенерируют и другие части тела, например конечности. Особенно наглядно эта способность выражена у личинок. Во взрослом состоянии она хорошо сохраняется у хвостатых; наоборот, бесхвостые после метаморфоза утрачивают ее почти вовсе.
Питание амфибий
Питание амфибий довольно однообразно. Пищей служат почти исключительно животные организмы, и только личинки кормятся и растениями. Бесхвостые и большинство хвостатых ловят главным образом беспозвоночных: земляных червей, пиявок, моллюсков и различных насекомых. Зеленые лягушки нападают иногда па птенцов, мелких грызунов и рыбу. Лягушка-бык и рогатая лягушка также ловят птенцов, мышей, крыс, иногда они хватают водяных змей. Гигантская саламандра питается в значительной мере рыбами и их икрой. Икру едят также и другие хвостатые. Безногие кормятся земляными червями и муравьями, в гнездах которых они нередко селятся.
Личинки большинства бесхвостых питаются в значительной мере растительной пищей. Они заглатывают плавающие водоросли и, кроме того, скоблят роговыми челюстями стебли подводных растений. Одновременно они захватывают и планктонных беспозвоночных: инфузорий, коловраток, мелких ракообразных. В связи с растительной пищей кишечник личинок бесхвостых относительно более длинный, чем у взрослых.
Личинки хвостатых и безногих питаются животными организмами.
Техника добывания пищи различна у разных групп. Бесхвостые ловят только двигающуюся добычу, которую они захватывают, выбрасывая клейкий язык. Реже (главным образом зеленые лягушки) они хватают добычу челюстями и лапами. Распознают добычу они, видимо, только зрением. Безногие и хвостатые отыскивают добычу и при помощи обоняния и ловят не только двигающиеся, но и неподвижные объекты, которые захватывают челюстями или языком.
Размножение
Внешние различия между самцами и самками у амфибий выражены в большинстве случаев сравнительно слабо. У некоторых (главным образом у бесхвостых) самцы несколько мельче самок. Самцы бурых лягушек весной имеют фиолетовый оттенок на спине и на горле. Яркая пятнистая окраска характерна для брачного наряда тритонов и протеев. Есть отличия и морфологического характера. Так, у самцов гребенчатого тритона весной кожистая фестончатая оторочка на спине и на хвосте разрастается особенно сильно, и в ней развивается густая сеть кровеносных сосудов. У самцов бесхвостых на внутренних пальцах передних ног имеются мозолистые утолщения, особенно развитые в брачный период, когда они помогают самцу плотнее обхватывать самку и крепче на ней держаться. У некоторых видов «брачные мозоли» имеются и на других частях конечностей.
Общей особенностью размножения амфибий является их связанность в этот период с водой в которую откладываются яйца и где происходит развитие личинок. Несомненно, что это было изначальной чертой земноводных. В последующем у ряда видов выработались новые приспособления, позволившие им размножаться и вне водоемов. Однако эти способности имеют явно вторичный характер, и они свойственны лишь немногим земноводным.
Оплодотворение у бесхвостых, за немногими исключениями, наружное. Самец, сидящий на спине самки, обхватывает ее передними лапами и сжимает с большой силой, выдавливая яйца. Объятия длятся долго, иногда несколько дней. Животные в это время держатся в воде. У большинства хвостатых и всех безногих оплодотворение внутреннее; однако только у последних бывает настоящее совокупление, и самцы у них имеют копулятивные органы. У тритонов самец после довольно длительного ухаживания за самкой выпускает в воду семя, заключенное в продолговатые пакетики — сперматофоры. Последние приклеиваются к подводным растениям, и самка, подплывая к ним захватывает их клоакой. У более наземных хвостатых (например, у Salamandra) спаривание бывает на суше. Самец обхватывает самку передними лапами, прикладывает свою клоаку к клоаке самки и выделяет в нее семя.
Оплодотворенные тем или иным путем яйца земноводных в подавляющем большинстве случа¬ев развиваются в водоемах, где они оставляются родителями на произвол судьбы. Наряду с этим известен ряд случаев, когда яйца развиваются в иной обстановке.
Безногие в большинстве случаев откладывают яйца в земляной норе, под корнями или под камнями. Отложив около 20 яиц, самка обвивается вокруг них и остается в таком положении до выхода личинок. Личинки (уже без наружных жабр) переселяются в воду, где очень быстро заканчивают метаморфоз.
У сумчатых лягушек (Nototrema) оплодотворенные яйца помещаются в глубокую складку кожи на спине. У некоторых видов головастики после выхода из сумки заканчивают свое развитие в воде, у других все стадии развития проходят в сумке, откуда выходят уже закончившие превращение лягушата.
Самки суринамской пипы (Pipa americana) откладывают яйца к себе на спину. Клоака к периоду размножения у них выпячивается, образуя длинный яйцеклад, который самец, влезая на самку, заворачивает ей на спину. Яйца вдавливаются самцом в сильно разбухающую к этому времени кожу спины самки так, что каждое яйцо располагается в своей ячейке. Число яиц в кладке бывает равным 50—100. Все стадии развития проходят в . ячейках, и наружу выходят лягушата, уже закончившие превращение.
В очень своеобразных условиях развиваются яйца южноамериканской лягушки — ринодермы Дарвина (Rhinoderma darwini). Самец заглатывает оплодотворенные яйца и проталкивает их в длинный голосовой мешок, расположенный под кожей на груди и брюхе. Одновременно в мешке помещаются 20 — 30 яиц. Развитие их только первоначально идет за счет желтка. В последующем личинки срастаются поверхностью спины и хвоста со стенками голосового мешка родителя и через богатую кровеносную сеть происходит обмен веществ. Наружу выходят лягушата, уже закончившие превращение.
Некоторые виды лягушек откладывают яйца в своего рода гнезда. Так, южноамериканские филломедузы спариваются на ветвях растений, свисающих над водой. Лягушки сближают края листьев и помещают в образовавшийся пакет оплодотворенные яйца. При этом края листьев склеиваются студенистыми оболочками яиц. Выведшиеся личинки некоторое время находятся в разжиженной массе оболочек, после чего вываливаются в воду, где и заканчивают метаморфоз. Другие виды тропических лесных лягушек откладывают яйца в пазухи листьев или приклеивают их к поверхности листьев, свисающих над водой. В последнем случае кладка яиц имеет вид комка, окруженного пенистой массой. На известных стадиях личинки переходят в воду.
Остается упомянуть оживородности у амфибий. Этот тип приспособления свойствен немногим из них и наблюдается чаще у хвостатых, реже у безногих и немногих видов пустынных жаб.
Так, пятнистая саламандра (Salamandra salamandra) обычно живородяща, и только иногда (при содержании в неволе) она откладывает яйца, из которых тотчас же выходят личинки, снабженные уже наружными жабрами.
У горной саламандры (Salamandra atra) весь метаморфоз происходит внутри тела матери. Вполне сформировавшихся детенышей рождает пещерная саламандра (Spelerpes fuscus) и некоторые виды безногих.
Все описанные случаи отклонения от нормального для амфибий типа размножения представляют приспособления, обеспечивающие успех воспроизведения видов, обитающих в своеобразной жизненной обстановке. Напомним, что внутреннее оплодотворение, «забота о потомстве», предохраняющая яйца от высыхания, или живорождение свойственны безногим, обитающим в совершенно необычной обстановке — под землей. В менее пригодных для развития яиц водоемах — в быстрых горных ручьях—возникает живорождение у некоторых саламандр. Наряду с этим многие виды их ведут преимущественно наземный образ жизни, и внутреннее оплодотворение в этих условиях, несомненно, является приспособительной особенностью.
Далее обращает внимание, что такие приспособления, как вынашивание яиц в кожистой сумке (нототрема), в голосовых мешках (ринодерма), устройство гнезд (филломедуза), присасывание личинок к телу взрослых при их миграциях (древолазы), свойственны видам, населяющим тропические об-ласти с их резкими чередованиями засушливых и дождливых периодов, и видам древесным, т. е. заселившим новую, вторичную для земноводных среду. Появление своеобразных приспособлений к размножению в этих необычных условиях вполне понятно.
Обращает внимание зависимость между совершенством приспособлений к размножению и плодовитостью. Так, черная саламандра родит единовременно обычно только двух детенышей, единичных детенышей рождают пятнистая саламандра и кольчатая череяга. Несколько большее число яиц имеют виды, вынашивающие их на теле или обвивающиеся вокруг яиц. Так, цейлонский рыбозмей откладывает 10—15 яиц, кольчатая череяга — 5—10 яиц, у пипы число яиц равно 50—100. у ринодермы Дарвина — 20—30. Еще больше яиц у видов, строящих гнезда для яиц: у летающей яванской лягушки — около 70, у филломедузы — около 100. Наконец, наибольшее число яиц откладывают виды, мечущие их в воду и оставляющие их на произвол судьбы. Травяная лягушка откладывает 1500—3000 яиц, зеленая лягушка — 3—8 тыс. икринок, жаба — около 10 тыс.
Развитие
Желток в яйцах амфибий распределен неравномерно и сконцентрирован на нижней, более светлой части яйца. Противоположная (анимальная) часть яйца черного цвета. Пигментированная шапочка на верхней анимальной части яйца, видимо, является приспособлением для защиты от вредного действия ультрафиолетовых лучей солнечного света. Яйцо испытывает полное, но неравномерное дробление. Через 8—10 суток (у лягушек) после оплодотворения зародыш прорывает яйцевые оболочки и в виде ли-чинки — головастика — выходит наружу. Первоначально у головастика нет парных конечностей и органом движения служит хвост, окаймленный хорошо развитой перепонкой. Первыми закладываются передние конечно¬сти, но они снаружи долго незаметны и сначала наружу появляются задние конечности. Вслед за этим у бесхвостых начинает укорачиваться, а затем и вовсе пропадает хвост.
Все личинки имеют в той или иной мере развитую боковую линию которая у безногих и бесхвостых (за немногими исключениями) затем исчезает. Органами дыхания первоначально служат две-три пары наружных ветвистых жабр. Далее у большинства видов наружные жабры атрофируются, и к этому времени развиваются жаберные щели с лепестками. Только у некоторых хвостатых наружные жабры сохраняются пожизненно. На этой стадии головастик имеет двухкамерное сердце, так как предсердие еще не поделено на левую и правую половины. Существует только один круг кровообращения, принципиально неотличимый от такового рыб. В последующем из переднего (глоточного) отдела пищеварительной трубки путем парных выпячиваний формируются легкие. Они снабжаются кровью от четвертой артериальной дуги, так как четвертая пара жабр у земноводных не развивается. По мере развития легких исчезают и внутренние жабры, а соответственно видоизменяется и кровеносная система. Первая пара жаберных сосудов превращается в сонные артерии, вторая — в системные дуги, третья у большинства не¬доразвивается, четвертая остается легочными артериями.
Наряду со сказанным происходит изменение и в других системах. Пронефрические почки исчезают и заменяются мезонефрическими. Кишечник укорачивается, исчезает боковая линия. Головастик становится лягушонком. Растительная пища, свойственная головастику, заменяется животной.
Неотения
Уже давно было замечено, что головастики некоторых бесхвостых (лягушек, жаб, чесночниц, жерлянок) не заканчивают в обычный срок метаморфоза и остаются личинками и на следующий год жизни. Рост их при этом не прекращается, и личинки достигают необычно больших размеров. Однако способности размножаться они не получают. Такое явление получило название неполной неотении. Более своеобразные уклонения в развитии свойственны хвостатым. У некоторых видов их личинки также достигают размеров взрослых (а иногда и более), у них созревают половые продукты, и животные начинают размножаться, сохранив все внешние личиночные признаки. Подобное явление, известное под наименованием полной неотении, свойственно некоторым видам тритонов и особенно американской амбистоме, неотеническая форма которой широко известна под именем аксолотля. Последнюю ученые долгое время считали за самостоятельный вид амфибий. В естественных условиях известны случаи нормального развития амбистомы, когда животное, закончив превращение, после утраты наружных жабр и после перехода от водного к наземному образу жизни начинает размножаться. Однако чаще и в природной обстановке встречаются неотенические формы амбистом, т. е. аксолотли.
При содержании в неволе превращение аксолотлей в амбистом можно получить путем соответствующих воздействий на развивающийся организм. Установлено, что у аксолотлей недоразвита щитовидная железа. Пониженная продукция ее и служит основной причиной неотении. При введении в организм аксолотлей гормона щитовидной железы (тироксина) начинается быстрый метаморфоз. Превращению способствует также изменение условий жизни. Затрудняя аксолотлям пребывание в воде и дыхание жабрами и стимулируя, наоборот, наземное существование и легочное дыхание, иногда удается вызвать метаморфоз. Механизм его, видимо, состоит в том, что при новых условиях жизни повышается активность щитовидной железы, назначение секрета которой для превращения уже указано выше. Надо думать, что и в природной обстановке полное превращение бывает в таких случаях, когда условия жизни обеспечивают повышенную продукцию щитовидной железы. Это связано, видимо, с особенностями питания, температурными условиями и влажностью.
Годовой цикл
Сравнительно малая приспособленность амфибий к обитанию в наземной среде обусловливает резкие изменения их образа жизни в связи с сезонными изменениями условий существования. Наиболее благоприятны они в экваториальной зоне. В густых тропических лесах относительно ровная температура и высокая влажность в течение всего года обеспечивают круглогодичное активное существование ряда амфибий. Однако в некоторых областях тропической зоны чередование дождливых и засушливых времен года приводит к резким изменениям условий жизни. В периоды засух амфибии исчезают. Они зарываются в ил, прячутся в норы, под корни или под камни. Так, например, на Яве такая летняя спячка длится около пяти месяцев.
В умеренных и северных широтах летней спячки не бывает и амфибии перестают быть активными зимой. Условия зимней спячки у разных групп амфибий различны. Наши лягушки собираются на места зимовок уже в сентябре, когда температура воздуха в среднем равна 8—12° С, а минимальная температура падает до 3—5° С. Отдельные взрослые особи встречаются под Москвой еще и в половине октября.
Вопреки распространенному мнению, бурые лягушки (R. temporaria) зимой обычно не закапываются в ил, а держатся на дне под камнями, в зарослях водорослей. Для зимовки выбираются ручейки с быстрым течением и с глубокими бочагами. В глубоких канавах и ямах с торфяным дном лягушки зимуют реже, и здесь они закапываются довольно глубоко в ил.
Жабы и квакши, наоборот, зимуют на суше в нежилых норах грызунов, под корнями деревьев, под камнями, в подвалах под домами.
Тритоны и саламандры зимуют также на суше. Они зарываются в мох, залезают в норы, под корни деревьев или под камни, где нередко собирается по нескольку десятков экземпляров. Установлено, что тритоны переносят более низкие температуры, чем лягушки. Они способны выносить охлаждение до —1,5° С, в то время как лягушки переносят охлаждение только до —0,5; —0,8° С.
Весной ранее всего пробуждаются бурые травяные лягушки. Под Москвой они появляются в первых числах апреля, иногда уже в конце марта. Зеленые лягушки выходят позднее, в первой половине мая. Тритоны пробуждаются тоже довольно рано, в половине апреля, когда местами (под Москвой) еще лежит снег.
После весеннего пробуждения амфибии ведут наиболее активный образ жизни, что связано с периодом размножения. Подавляющее большинство их переселяется в воду, где происходит спаривание и откладывание икры. После размножения поведение существенно меняется. Жабы и травяные лягушки покидают водоемы. Жабы становятся активными только ночью, травяные лягушки также активны преимущественно в сумерках и ночью. Тритоны остаются в воде около 2—3 месяцев, и в это время они активны круглые сутки. В половине лета тритоны покидают водоемы и становятся сухопутными животными. В этот период они активны только ночью.
Укажем, что в связи со сменой условий жизни меняется и организация животных. Так, у тритонов в период водного существования кожа тонкая, обильно увлажняемая выделениями желез. При жизни на суше кожа становится толстой и грубой, что, несомненно, является приспособлением к жизни вне воды. Одновременно меняется и внешний вид животного. У обыкновенного тритона вовсе исчезает бахромчатый плавник, у гребенчатого тритона он заметно уменьшается. Установлено, что разрастающаяся в период водного существования кожистая оторочка на спине и на хвосте имеет прямое приспособительное значение. Тритоны в это время дышат в значительной мере через кожу, и указанные оторочки служат местом наибольшего ветвления кожных кровеносных сосудов.
Осенью начинается миграция амфибий к местам зимовок. Особенно заметны передвижения травяных лягушек, которые в это время собираются в довольно значительные группы. Меняется и их суточная активность. Из ночных они становятся дневными, что связано с понижением температуры ночью. Уже при 8—10° С активность лягушек заметно снижается. Надо учесть, что понижение температуры обусловливает прекращение активности насекомых и червей, служащих лягушкам кормом.
Таким образом, сезонные изменения условий жизни вызывают резкие изменения биологии и условий нахождения амфибий.
Значение земноводных
Сравнительно с другими позвоночными земноводные имеют небольшое практическое значение. В общем земноводные полезны для человека. Лягушки и жабы уничтожают много вредных беспозвоночных животных: членистоногих, моллюсков. Водные виды поедают личинок вредных насекомых и животных, распространяющих болезни. Так, тритоны уничтожают личинок комаров, в том числе и тех их видов, которые передают малярию. Польза амфибий, особенно жаб, усиливается тем, что они часто активны ночью, когда спит большинство насекомоядных птиц.
По подсчетам, произведенным Красновидовым в Новгородской области, одна травяная лягушка съедает за сутки в среднем 7 вредных животных (насекомых, их личинок, брюхоногих моллюсков), а за 6 месяцев периода бодрствования — около 1200. Было установлено, что на площади в 24 тыс. м2 обитало 720 особей лягушек, которые, согласно этим расчетам, вероятно, уничтожили около 900 тыс. экземпляров вредных животных.
Некоторые амфибии служат пищей для ценных пушных хищников, например для черного хоря и енотовидной собаки, пища которой состоит местами более чем наполовину (до 65%) из лягушек и головастиков. Лягушками и головастиками кормятся и многие полезные птицы, например: утки, журавли и др. Есть опыты разведения головастиков для откорма домашних уток. Наконец, в некоторых странах едят мясо лягушек и крупных саламандр.
В ряде стран добыча амфибий без специальных разрешений запрещена.
Известны случаи преднамеренного завоза и выпуска их (главным образом жаб) для биологической борьбы с вредителями сельского хозяйства.
Надо учесть, что лягушек в громадном количестве используют для научных и учебных целей.
Реальная отрицательная роль амфибий практически не установлена. Номинально она проявляется лишь местами, в определенных условиях. Так, озерная зеленая лягушка поедает молодь рыб. В желудках лягушек, добытых в местах скопления молоди рыб, находили до 30—40 мальков сазана и воблы длиной 10—20 мм. Однако исследования В. К. Маркузе (1964), проведенные в низовьях Волги в условиях нерестовыростных хозяйств, показали, что вся популяция лягушек за время нахождения мальков на рыбхозе уничтожает менее 0,1 % их. Неясно значение лягушек при прудовом разведении карпа. Хотя они уничтожают его мальков, но размер изъятия не выяснен.
Поделиться с друзьями