
ISSN 0869-5938, Stratigraphy and Geological Correlation, 2007, Vol. 15, No. 3, pp. 310-320. © Pleiades Publishing, Ltd., 2007.
Original Russian Text © L.G. Bragina, Yu.V Agarkov, N.Yu. Bragin, 2007, published in Stratigrafiya. Geologicheskaya Korrelyatsiya, 2007, Vol. 15, No. 3, pp. 76-86.
Radiolarians of the Upper Cenomanian and Lower Turonian from Deposits of the Ananuri Formation, the Western Caucasus (Lazarevskoe Area)
L. G. Bragina", Yu. V. Agarkov*, and N. Yu. Bragin"
a Geological Institute, Russian Academy of Sciences, Moscow, Russia b Rostov State University, Rostov-on-Don, Russia Received May 16, 2005; in final form, September 29, 2006
Abstract—Diverse radiolarians (over 70 species) are detected in cherty rocks above the bituminous shale hori¬zon, the marker of anoxic event OAE-2 recorded across the Cenomanian-Turonian boundary in the upper part of the Ananuri Formation of flyschoid deposits, the Lazarevskoe area of the western Caucasus. The radiolarian assemblages studied are comparable in composition with radiolarians from concurrent Cenomanian-Turonian boundary strata in other Mediterranean regions (e.g., in the Crimea and Turkey). The lower radiolarian assem¬blage includes index species Dactyliosphaera silviae of synonymous Cenomanian zone. Alievium superbum present in the upper assemblage is index species of the relevant Turonian zone. Within the studied flyschoid sequence, sediments indicative of the above event (bituminous shales and cherts) are confined to upper elements of flysch rhythms.
DOI: 10.1134/S0869593807030069
Key words: western Caucasus, Cenomanian, Turonian, radiolarians, anoxic events, flysch.
INTRODUCTION
Radiolarians of the lower Upper Cretaceous in Tethyan areas (Italy and Spain) have been studied in detail during the last two decades (Erbacher, 1994; O’Dogherty, 1994; Salvini and Marcucci Passerini, 1998). Of special importance is monograph by O’Dogherty (1994) who described dozens of new taxa and revised species originally identified by Squinabol (1903, 1904, 1914). In radiolarian zonation suggested by O’Dogherty for the Barremian-lower Turonian interval, radiolarian subdivisions in the Cenomanian- Turonian transition, where the Oceanic Anoxic Event (OAE-2) of global extent is recorded, are of particular interest (Kuhnt et al., 1986; Schlanger et al., 1987). It is confidently established that this event greatly influ¬enced composition of biota (Vishnevskaya et al., 2006), in particular, of radiolarians (Bragina, 2001). In Italy and Spain, the event is recorded in the so-called “Bonarelli Beds” with elevated content of organic car¬bon, which represent a reliable regional reference hori¬zon. The Cenomanian-Turonian boundary is defined at the Alievium superbum Zone base exactly in the “Bonarelli Beds.” As is established, termination of the OAE-2 was followed in different regions of the world by quantitative increase of radiolarian productivity, tax¬onomic diversification of radiolarian assemblages, and by deposition of siliceous radiolarian sediments (O’Dogherty, 1994; Bragina, 1999, 2001).
Bragina (2004) who studied Cenomanian-Turonian radiolarians in the Crimean Mountains and northern Turkey verified taxonomic composition of their assem¬blages and stratigraphic ranges of certain species. Hav¬ing distinguished the relevant radiolarian zones in these regions, she failed to establish here subdivisions of sub¬zone rank. It can be assumed therefore that index spe¬cies of radiolarian subzones recognized in Italy and Spain are of a limited geographic range, but the assumption should be checked on radiolarian succes¬sions in other regions. Accordingly, we carried out fieldwork in the Lazarevskoe area of the western Cau¬casus, where the widespread Upper Cretaceous sequence of deep-water flysch contains radiolarians at different stratigraphic levels. Our attention was concen-trated on the Ananuri Formation composed in its upper part of clay and chert beds with interlayers of bitumi¬nous shales, the OAE-2 markers. The overlying Kerket Formation is composed in lower interval of flaggy lime¬stone beds with interlayers of radiolarian chert. During fieldwork, we discovered abundant and diverse radi¬olarian assemblages confined to chert interlayers near the Ananuri Formation top. These assemblages are very suitable for comparison with concurrent assemblages of the Crimea and other Mediterranean regions.
Radiolarians of the Greater Caucasus attracted attention of geologists since the 1930s (Vyalov, 1934; Karstens, 1932). To a considerable extent, these are Cenomanian-Turonian radiolarians of the Ananuri chert horizon, which extends along southern flank of the Greater Caucasus from Tuapse across Abkhasia, Kutaisi region and South Osetia to Ananuri and further via Kakhetia to southeastern Azerbaijan (Vishnevskaya, et al., 1990, 2006).
In the Greater Caucasus western flank, radiolarians have been studied earlier at the base of carbonate sequence exposed along the Ol’khovaya and Khosta rivers, where they are confined to cherts and bitumi¬nous marls of the Ananuri Horizon (Vishnevskaya, 2001). Radiolarians confined to topmost horizons of the Ananuri Formation in the Lazarevskoe section have not been studied until this work. We macerated radiolarians from cherts sampled here using conventional method of rocks decomposition in 5% solution of hydrofluoric acid (Nazarov and Vitukhin, 1981).
SECTIONS OF ANANURI FORMATION AND DISTRIBUTION OF RADIOLARIANS
The thick carbonate-terrigenous flysch of the Upper Cretaceous is a remarkable sedimentary sequence in southern flank of the western Caucasus. Its Cenoma¬nian-Turonian interval is divided here (Agarkov et al., 1992; Afanas’ev and Maslakova, 1967; Vassoevich, 1947; Keller, 1940) into the Pauk (Cenomanian sand¬stones, siltstones, shales and tuffites), Ananuri (Cenom- anian-lower Turonian cherty limestones, cherts, marls, sandstones, siltstones and combustible shales), Kerket (lower Turonian cherty limestones and cherts), and Natukhai (upper Turonian-Coniacian limestones, marls and clays) formations. We studied two sections, one in outskirts of the settlement Lazarevskoe, the Sochi district, and the other one near the settlement Volkonka, the Mamedova Shchel Canyon (Fig. 1), which are similar to each other. Samples for micropal- eontological analysis have been collected from upper elements of flyschoid rhythms, i.e., from most fine¬grained lithologic varieties of sediments. Depending on the rhythm thickness, we collected three to six samples per one meter of the section.
In a railway excavation to the northwest of the Station Volkonka, there is exposed the rock succession of the Pauk, Ananuri, and Kerket formations. The upper part of the Ana- nuri Formation is here of the following structure (from the base upward):
(1) Rhythmically alternating sandstones, siltstones, and shales. Gray to greenish gray, medium- and fine-grained sandstones with horizontal, sometimes wavy and cross strat¬ification contain plant detritus confined to bedding planes, the basal of which are decorated with hieroglyphs (individual layers from 1 to 10 cm thick). In gray siltstones with con- choidal fracture, very fine horizontal stratification is empha¬sized by light-colored sandy laminae. Gray to dark greenish gray shales are calcareous sometimes. In addition, we observed rare beds of gray marl and single layers of black combustible shales foliated and ferruginate. Apparent thick¬ness is 3.7 m, strike NW 290°, dip 30°.
(2) Rhythmical interlayering of sandstones, siltstones, shales, combustible shales, and cherts. Gray to brownish gray
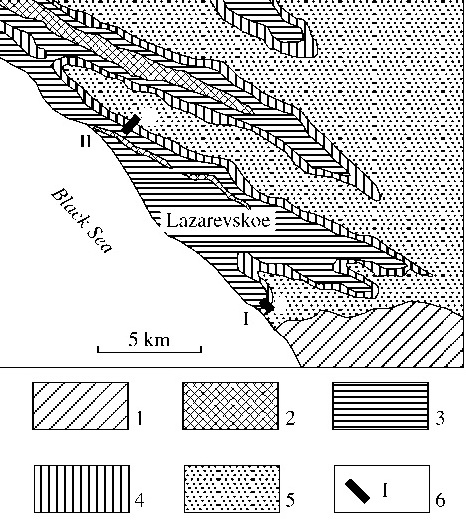
Fig. 1. Geologic scheme of the Lazarevskoe area, southern flank of the western Caucasus: (1) Maastrichtian, (2) Cam¬panian, (3) upper Turonian-Santonian, (4) upper Cenoma- nian-lower Turonian (Ananuri and Kerket formations), (5) Cenomanian and Lower Cretaceous deposits; (6) locali¬ties of the Volkonka (I) and Mamedova Shchel (II) sections.
fine-grained sandstones with thin horizontal stratification (3¬6 cm) contain small-sized plant detritus at bedding planes. Fine horizontal lamination in gray siltstones (5-15 cm) sometimes grades into vague cross and wavy lamination. Gray to dark and greenish gray laminated shales (1-5 cm) are siliceous in places. Black flaggy laminated combustible shales (2-8 cm) are ferruginate, rusty when weathered, con-taining fine laminae composed of sulfides and jarosite, pan-cake-shaped pyrite nodules and dispersed gypsum crystals and rosettes. Their bedding planes are smeared with easily inflammable bitumen. Greenish gray clayey shales (2-5 cm) commonly grade into siliceous shales. Total thickness is 1.5 m.
(3) Rhythmical interlayering of siltstones, shales and cherts. Gray to yellowish gray siltstones (2-3 cm) have very fine horizontal lamination. Greenish to yellowish gray shales are foliated (laminae 0.5 cm thick). In radiolarian greenish gray, light yellowish gray and white flaggy cherts clayey sometimes (4-14 cm), fine one-mm-thick horizontal lamina-tion is emphasized by laminae with concentrated radiolarians and dispersed pyrite frequently replacing microfossils. Radi- olarians described below are found at two levels: at the level of 45 cm above the bed base (Sample L-2-3) they belong to the Dactyliosphaera silviae assemblage and at the higher level (60 cm above the base) to the Alievium superbum assemblage. Apparent thickness is 0.65 m.
Chocolate brown marls of the Kerket Formation appearing after unexposed section interval 0.5 m thick grade upward into red, pink and light yellow-gray cherty limestones and marls
(Fig. 2).
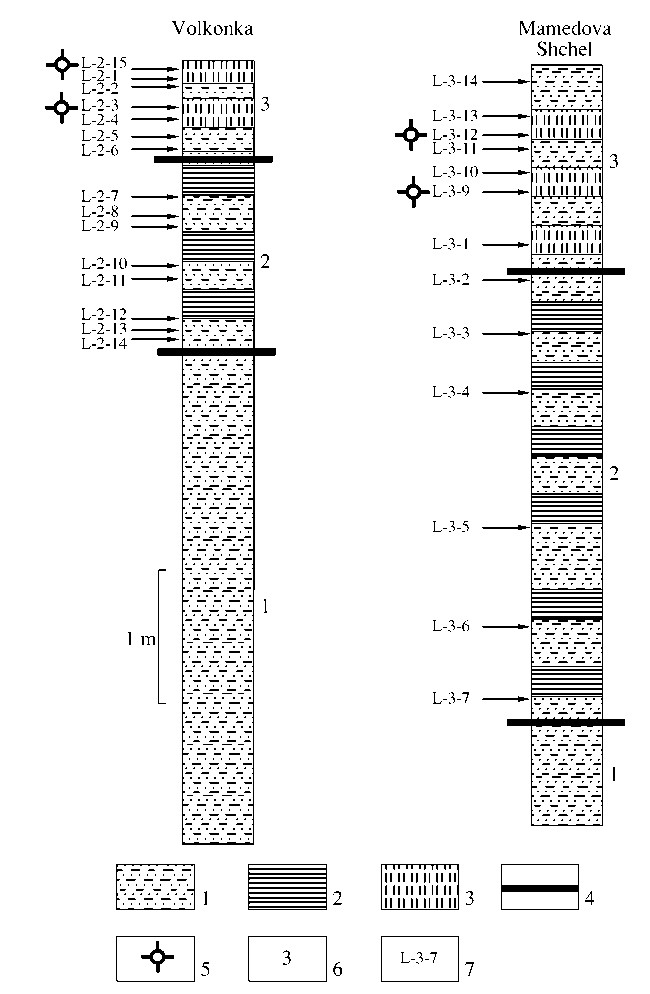
Fig. 2. Lithologic structure of the Cenomanian-Turonian transition in the Volkonka and Mamedova Shchel sections: (1) sandstones, siltstones and shales; (2) combustible shales; (3) cherts; (4) boundaries between beds described in the text; (5) occurrence levels of radiolarians; (6) bed numbers; (7) sample numbers.
The Dactyliosphaera silviae assemblage includes the following species: Acaeniotyle diaphorogona Fore¬man, Acanthocircus impolitus O’Dogherty, A. tympa¬num O’Dogherty, A. sculptus (Squinabol), Archaeo- cenosphaera ? mellifera O’Dogherty, Cavaspongia antelopensis Pessagno, C. californiaensis Pessagno, Crucella aster (Lipman), C. messinae Pessagno, Dac- tyliodiscus lenticulatus (Jud), D. longispinus (Squin¬abol), Halesium quadratum Pessagno, H. sexangulum Pessagno, Orbiculiforma ovoidea Bragina, Paronaella spica Bragina, Patulibracchium ingens (Lipman), Pes- sagnobrachia irregularis (Squinabol), Phaseliforma inflata Bragina, Phaseliforma sp. ex gr. P. inflata Brag¬ina, Praeconocaryomma californiaensis Pessagno, P. lipmanae Pessagno, P. universa Pessagno, Pyrami- spongia glascockensis Pessagno, Quinquecapsularia ombonii (Squinabol), Stylodictya insignis Campbell et Clark, Triactoma cellulosa Foreman, Amphipyndax stocki (Campbell et Clark), Diacanthocapsa antiqua (Squinabol), D. euganea Squinabol, D. fossilis Squin¬abol, D. ovoidea Dumitrica, D. rara Squinabol, D. sp. A, Dictyomitra montisserei (Squinabol), Distylocapsa squama O’Dogherty, D. veneta (Squinabol), Holocryp- tocanium astiensis Pessagno, Phalangites telum
O’Dogherty, Pseudodictyomitra nakasekoi Taketani, P. pseudomacrocephala (Squinabol), Pseudoeucyrtis pulchra (Squinabol), Spongostichomitra elatica O’Dogherty, Stichomitra communis Squinabol, and Xitus spineus Pessagno.
None of the taxa obviously prevails over others in the assemblage, and preservation degree of radiolarians is of selective character. For instance, their delicate thin-walled tests are greatly fragmented being inappro¬priate for precise identification. Nevertheless, these clasts suggest indirectly a much higher taxonomic diversity of radiolarian community at the lifetime. We managed to identify 45 species of the Dactyliosphaera silviae assemblage characteristic of both the upper Cenomanian and lower Turonian deposits in different Tethyan regions, e.g., in Italy, Spain, and Turkey. The assemblage includes a series of species first described by O’Dogherty (1994) from Italy and Spain (Acantho- circus impolitus, A. tympanum, Archaeocenosphaera ? mellifera, Distylocapsa squama, Phalangites telum, Spongostichomitra elatica) and by Bragina (2004) from Turkey (Paronaella spica, Phaseliforma inflata, Orbiculiforma ovoidea), and their geographic ranges can be considered wider therefore.
In radiolarian zonation suggested by O’Dogherty (1994) for the Tethyan Superrealm, the Dactyl¬iosphaera silviae Zone of the terminal lower and upper Cenomanian is divided into the lower Patellula spica and upper Guttacapsa biacuta subzones. Zonal index species Dactyliosphaera silviae is present in the stud¬ied assemblage, but index species of two subzones have not been detected like in some other Tethyan regions, e.g., in northern Turkey and Crimean Mountains (Bra¬gina, 2004). An interesting feature of our assemblage is last occurrence of Spongostichomitra elatica, the spe¬cies not recorded above the lower Cenomanian in sec¬tions of Italy and Spain (O’Dogherty, 1994), in associ¬ation with Diacanthocapsa antiqua and Pseudoeucyrtis pulchra regarded to be very characteristic of the upper Cenomanian (O’Dogherty, 1994). Of particular interest is presence of Acanthocircus tympanum that appears in sections of Italy and Spain concurrently with Alievium superbum, the index taxon of the synonymous Turonian zone. Taking into account the established coexistence of Dactyliosphaera silviae, the index species of synon¬ymous Cenomanian zone, and Acanthocircus tympa¬num characteristic of the Turonian, it is possible to sug¬gest that the described assemblage corresponds in age to the terminal Cenomanian.
The Alievium superbum assemblage includes many species of the Dactyliosphaera silviae assemblage being simultaneously enriched in radiolarians, which do not occur at the lower level. These are Acaeniotyle macrospina (Squinabol), Alievium superbum (Squin¬abol), A. cf. sp. A. superbum (Squinabol), Cavaspongia contracta O’Dogherty, Crucella cachensis Pessagno, Patellula verteroensis (Pessagno), Pessagnobrachia rara (Squinabol), Phaseliforma subcarinata Pessagno,
RADIOLARIANS OF THE UPPER CENOMANIAN AND LOWER TURONIAN
Pseudoaulophacus floresensis Pessagno, P praeflore- sensis Pessagno, Triactoma compressa (Squinabol), T. fragilis Bragina, T. hexeris O’Dogherty, Vitorfus brus- tolensis (Squinabol), V morini Empson-Morin, Archaeodictyomitra sliteri (Squinabol), A. sp. ex gr. A. squinaboli Pessagno, Cryptamphorella conara (Foreman), C. sphaerica (White), Diacanthocapsa elon- gata Bragina, Holocryptocanium barbui Dumitrica, Phalangites hastatus O’Dogherty, Pogonias ? hirsutus (Squinabol), Squinabollum fossile (Squinabol), Sti- chomitra insignis Squinabol, S. magna Squinabol, Tha- narla sp. ex gr. T. conica (Aliev), Tubilustrium trans- montanum O’Dogherty, and Xitus spicularius (Aliev).
The assemblage under consideration consists of 73 species, the majority of which (60%) is known from Cenomanian and lower Turonian deposits of the Tethys. Nearly all the newcomers are also widespread in the same stratigraphic intervals. Radiolarians of the Alievium superbum assemblage are preserved rather well, as very fragile tests representing the genus Vitor¬fus are present among them. However, delicate thin- walled morphotypes are dilapidated and hardly identi¬fiable. The assemblage includes species Phaseliforma inflata and Diacanthocapsa elongata described from the Cenomanian of Turkey (Bragina, 2004), Triactoma fragilis known from the Santonian-Campanian of Cyprus (Bragina and Bragin, 1996) and found also in the Cenomanian of Turkey (Bragina, 2004). Species Paronaella spica occurs in the Cenomanian of northern Turkey and in the lower Turonian of the Crimean Mountains (Bragina, 2004). The age interpretation of the Alievium superbum assemblage is based on coexist¬ence of (1) Dactyliosphaera silviae, the index species of synonymous Cenomanian zone unknown from the Turonian of Italy and Spain, and (2) Alievium super¬bum, the index species of synonymous Turonian zone (Pessagno, 1976; O’Dogherty, 1994). Based on associ¬ation of these two taxa, we assume that the Alievium superbum assemblage corresponds in age to earliest Turonian time, when first species Alievium superbum appeared to join still existing Dactyliosphaera silviae forms.
At the Mamedova Shchel site 2.5 km upstream of the Kuapse River mouth, there are sporadic outcrops of the Aptian-Albian terrigenous deposits and overlying rocks of the Pauk Formation. The Ananuri Formation upper part and the entire Kerket Formation are observ¬able further upstream in a continuous exposure. Transi¬tion between the last two formations is of the following structure here (from the base upward, Fig. 2):
(1) Rhythmical interlayering of limestones, marls and shales. Greenish gray limestones (10-30 cm) are clay-cherty, aphanitic. Green to greenish gray layers of marl are thin (2¬15 cm). Gray to dark gray shales (2-6 cm) are foliated. Apparent thickness is 0.7 m.
(2) Rhythmical interlayering of sandstones, siltstones, shales and combustible shales. Beds of gray fine-grained sandstones (2-10 cm) with hieroglyphs at the base have hor¬izontal and more rare wavy and cross lamination. Dark gray,
gray and greenish gray siltstones and shales (1-15 cm) are thin-laminated, foliated, sometimes calcareous, grading into marls. Black combustible shales (1-5 cm) are flaggy, foli¬ated, containing jarosite at foliation planes. Thickness is 3.25 m.
(3) Rhythmical interlayering of sandstones, marls, lime¬stones and cherts. Gray fine-grained sandstones have fine horizontal stratification (0.5-1 cm). Gray to greenish gray marls (5-15 cm) contain fucoids. Greenish to yellowish gray limestones (2-15 cm) are cherty and flaggy. In greenish, yel¬lowish and reddish gray, banded radiolarian cherts (5-15 cm) with fine horizontal lamination, radiolarians are concentrated in separate laminae. Total thickness is 1.4 m.
The above beds 1-3 belong to the Ananuri Forma¬tion and are overlain by reddish, yellowish and greenish limestones and marls of the Kerket Formation. We stud¬ied radiolarians from two levels of Bed 3: 55 cm (Sam¬ple L-3-9) and 1 m (Sample L-3-12) above the bed base.
The radiolarian assemblages from the Mamedova Shchel section are less representative in general than their counterparts from the Volkonka section most likely due to a worse preservation of radiolarian skele¬tons. The lower assemblage (Sample L-3-9) includes Archaeocenosphaera ? mellifera O’Dogherty, Crucella irwini Pessagno, Godia coronata (Tumanda), Halesium sexangulum Pessagno, Patulibracchium woodlanden- sis Pessagno, Phaseliforma inflata Bragina, Praecono- caryomma lipmanae Pessagno, Protoxiphotractus cf. sp. P ventosus Pessagno, Pyramispongia glascockensis Pessagno, Cryptamphorella sphaerica (White), Dia- canthocapsa antiqua (Squinabol), D. elongata Brag¬ina, Distylocapsa squama O’Dogherty, Pseudodictyo- mitra pseudomacrocephala (Squinabol), Squinabollum fossile (Squinabol), Stichomitra communis Squinabol, and Xitus spicularius (Aliev). The assemblage includes 17 species, many of which are universally widespread in the Cenomanian of the Tethyan Superrealm. Diacan- thocapsa elongata Bragina that is present in the assem¬blage and has been originally described from Cenoma- nian deposits of northern Turkey (Bragina, 2004) extends geographic range of this species. The assem¬blage is lacking index species of Cenomanian and Turonian radiolarian zones, and its stratigraphic range cannot be defined precisely. Presumable age limits are the late Cenomanian and early Turonian, because the greater part of listed species is characteristic of both time spans.
The assemblage occurring 0.45 m higher in the sec¬tion includes nearly all the species of previous assem¬blage. Species unknown below and first occurring at this level are Acanthocircus tympanum O’Dogherty, Alievium sculptus (Squinabol), A. sp. cf. A. superbum (Squinabol), Archaeospongoprunum sp. A, Cavaspongia antelopensis Pessagno, C. californiaensis Pessagno, C. euganea (Squinabol), Crucella cachensis Pessagno,
C. messinae Pessagno, C. aff. C. messinae Pessagno, Patellula verteroensis (Pessagno), Pessagnobracchia irregularis (Squinabol), Phaseliforma sp. ex gr. P inflata Bragina, Praeconocaryomma sp. ex gr. P lipmanae Pessagno, Pseudoacanthosphaera galeata O’Dogh- erty, Pseudoaulophacus praefloresensis Pessagno, P. putahensis Pessagno, Amphipyndax stocki (Camp¬bell et Clark), Diacanthocapsa sp. ex gr. D. antiqua (Squinabol), D. brevithorax Dumitrica, D. sp. ex gr.
D. elongata Bragina, D. cf. ex gr. D. fossilis (Squin- abol), Dictyomitra sp. ex gr. D. densicostata Pessagno, Holocryptocanium barbui Dumitrica, and Stichomitra insignis (Squinabol). The assemblage includes 43 spe¬cies in total, but well-preserved specimens of Alievium superbum have not been detected, and we managed to identify only Alievium cf. sp. A. superbum. As Acantho- circus tympanum is found in association with species widespread in the Cenomanian-Turonian transition, their host deposits span the late Cenomanian-early Turonian interval in our interpretation.
Thus, the described radiolarian assemblages surely prove the upper Cenomanian-lower Turonian strati- graphic range of the Ananuri Formation upper part, and relevant combustible shales can be interpreted as sedi¬ments of the OAE-2. The last inference is important for comparative analysis of this event recorded in different sections and facies of the Mediterranean region.
Sections with the OAE-2 records are comprehen¬sively studied in Italy, where this event took place in a relatively deep basin with slow hemipelagic sedimenta¬tion in quiet environments (O’Dogherty, 1994). In upper part of the Scaglia Bianca Formation (middle Albian-lower Turonian) represented by fairly thin (65¬70 m) sequence of alternating micritic limestones and cherty rocks, there have been distinguished the so- called “Bonarelli Beds” with interlayers of bituminous shales most carefully studied in the Gorgo a Cerbara, Gola del Bottaccione, and Monte Casalini sections of the Umbria-Marche Apennines (O’Dogherty, 1994). The Bonarelli Beds are of the following structure in these sections: (1) lower interval (30-100 cm) of light- colored siliceous shales, siltstones and radiolarites with minor admixture of organic matter; (2) middle interval (30-100 cm) of black shales enriched in organic matter and phosphorus, frequently containing fish remains;
Radiolarians of the Cenomanian-Turonian transition in the Volkonka section, southern flank of the western Caucasus (Volkonka section, plates 1, 2).
Plate I. (1) Acanthocircus tympanum O’Dogherty, x200; (2) Acanthocircus impolitus O’Dogherty, x200; (3) Archaeocenos¬phaera ? mellifera O’Dogherty, x150; (4, 5) Acaeniotyle macrospina (Squinabol), x300; (6) Phaseliforma sp. ex gr. P inflata Bra¬gina, x150; (7) Multastrum sp. ex gr. M. regalis Vishnevskaya, x200; (8) Pseudoacanthosphaera sp. ex gr. P magnifica (Squinabol), x100; (9) Pessagnobracchia irregularis (Squinabol), x150; (10) Crucella messinae Pessagno, x200. Figs. (1, 4, 5, 9) - Turonian; fig. (10) - upper Cenomanian.
(3) upper interval (10-30 cm) of radiolarian shales, silt- stones and cherts. Within the last interval, concentra¬tion of organic matter declines gradually upward, while content of carbonate components increases.
A similar horizon corresponding to the OAE-2 is known in sections of the southwestern Crimea (Naidin and Alekseev, 1980; Naidin et al., 1981; Naidin and Kiyashko, 1994a, 1994b), which are composed of alter-nating limestone and marl beds. A horizon of black bituminous marls, which contain fish remains and later-ally grade into marls with elevated content of organic matter, is locally observable here at the level of the Cen- omanian-Turonian boundary. In sections of the Mt. Selbukhra and Mt. Belaya, this horizon is overlain by bed (20-30 cm) of cherty and highly silicified lime-stones with radiolarians. Despite certain distinctions in structure and lithologic composition, sections of the Crimea and Italy reveal an important feature in com-mon: bituminous shales grade upward into cherty sedi-ments with radiolarians crowned by carbonate deposits.
In southern flank of the western Caucasus, sections of the Ananuri Formation have been deposited in deep- water settings under conditions of intense influx of clastic material (Afanas’ev, 2004). Sedimentation rate was much higher here than in the southwestern Crimea. In the Volkonka and Mamedova Shchel sections, the OAE-2 is recorded against background of concurrent flyschoid sedimentation. As a result, we observe here not one but several horizons of combustible shales rep-resenting crowning elements D or E in typical rhythms of turbidity flysch (Bouma, 1962) that characterizes completely or partially the background sedimentation in the basin. The lower elements of rhythms (A, B, C) composed of sandstones with graded bedding and fine-grained siltstones are lacking signs of the OAE-2, although it is clear that this event was concurrent to but independent of the turbidity sedimentation. In a similar manner, radiolarian cherts appearing above last hori¬zons of combustible shales are confined to terminal ele¬ments of flyschoid rhythms. As a consequence, total thickness of sediments deposited at the OAE-2 time in southern flank of the western Caucasus is essentially greater than in Italy, for instance, and their lithologic composition is much more diverse because of deposi¬tion of siliciclastic material. Nevertheless, the general trend of sedimentogenesis first established in the Bonarelli Beds and corresponding to lithologic succes¬sion of bituminous shales, radiolarian cherts and car¬bonate rocks almost lacking organic matter is traceable in the studied sections as well. Accordingly, the anoxic event (OAE-2) was similarly manifested in different settings of deep-sea basin: in a zone of hemipelagic or even pelagic sedimentation (Italy), shelf zone (Crimea), and a relatively deep flysch trough (southern flank of the western Caucasus).
Data on taxonomic diversity of radiolarian assem-blages from different sections of the Tethys elucidates paleogeography of these fossil organisms. There is no doubt of course that Tethyan basins have been largely populated by identical radiolarian taxa. Nevertheless, it is possible to state even now that concurrent radiolarian assemblages were dissimilar to a certain extent. For example, comprehensive study of the middle-upper Cenomanian radiolarians in northern Turkey (Bragina, 2004) showed that 32 species of their assemblages are unknown in concurrent deposits of Italy and Spain. Pre-liminary data on the Cenomanian-Turonian radiolari¬ans from the Perapedhi and Moni formations of Cyprus (Bragina and Bragin, 2006) also imply that some of their species do not occur in the Crimean Mountains, Turkey (Bragina, 2004), Italy and Spain (Erbacher, 1994; O’Dogherty, 1994; Salvini and Marcucci Passe- rini, 1998). In this work, some radiolarian taxa from the southwestern Caucasus are identified in open nomen-clature and need subsequent systematic description.
Despite the taxonomic diversity of upper Cenoma- nian-lower Turonian radiolarians in the study objects, we failed to detect among them the index species of the Guttacapsa biacuta Subzone (established by O’Dogh- erty in the upper Cenomanian of Italy and Spain) in either the Volkonka or the Mamedova Shchel sections. The studied assemblages are lacking also some other species (Sciadiocapsa pertica, Diacanthocapsa matsu- motoi, Dictyomitra crassispina) typical of this subzone. As Guttacapsa biacuta is unknown as well in the Crimean Mountains and northern Turkey, we suspect that the relevant subzone is of a limited geographic range.
SOME CHARACTERISTIC RADIOLARIAN SPECIES
Acanthocircus tympanum O’Dogherty, 1994, emend. Bragina, 2004
Plate I, fig. 1
Mesosaturnalis sp.: Erbacher, 1994, pl. 19, figs. 6, 7.
Acanthocircus tympanum: O’Dogherty, 1994, p. 259, pl. 45, figs. 17-24; Salvini and Marcucci Passerini, 1998, Fig. 8.p; Bragina, 2004, pl. 34, fig. 15.
Distribution: lower Turonian of Italy and Spain; lower Turonian of the Crimean Mountains; uppermost Cenomanian-basal Turonian of the western Caucasus.
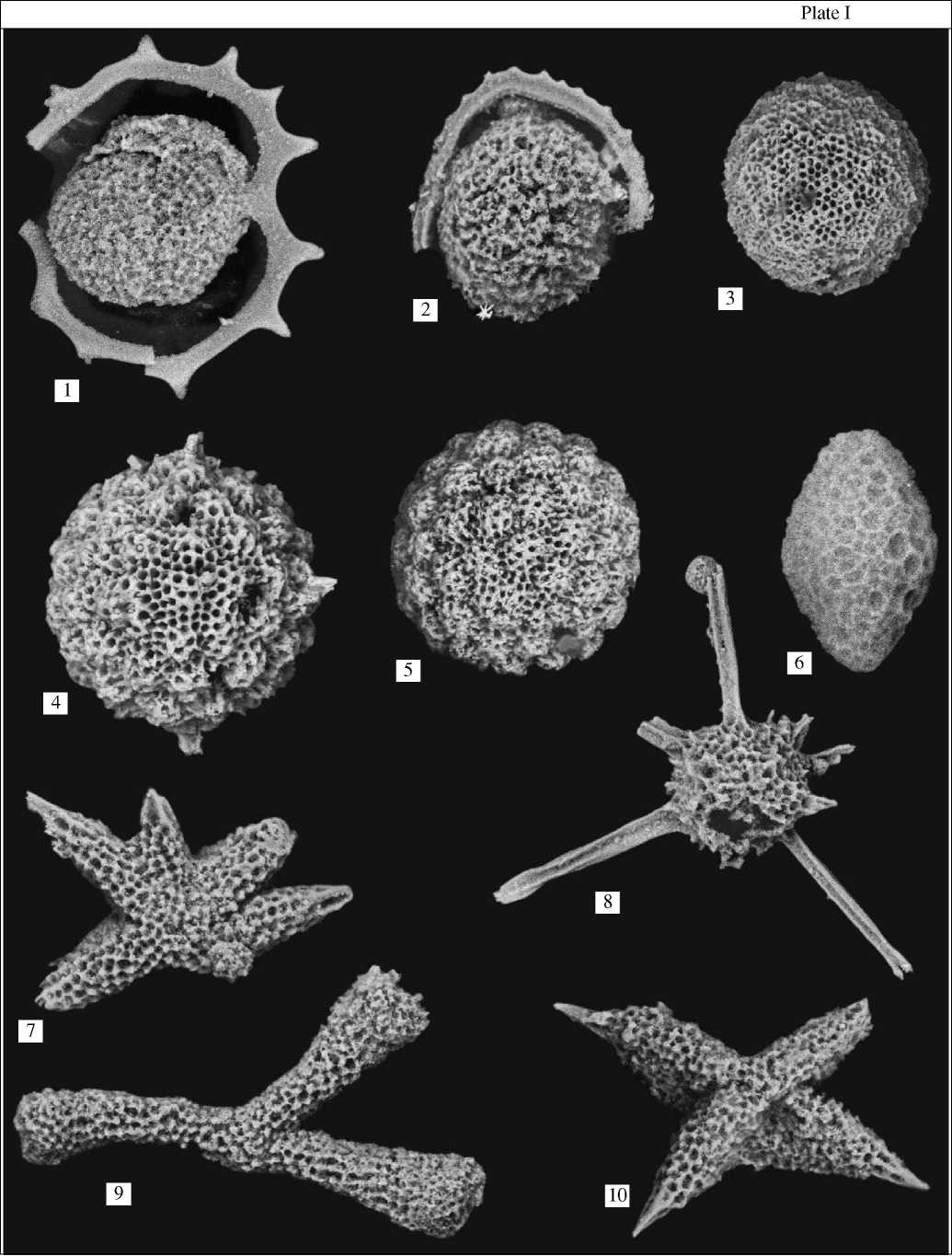
Plate II. (1) Pseudoaulophacus praefloresensis Pessagno, x200; (2) Alievium cf. sp. A. superbum (Squinabol), x200; (2) Cava- spongia californiaensis Pessagno, x200; (3) Crucella messinae Pessagno, x250; (5, 6) Dictyomitra cf. sp. D. multicostata Zittel, x120 (both); (7) Pseudodictyomitra pseudomacrocephala (Squinabol), x150; (8) Diacanthocapsa euganea Squinabol, x200; (9) Diacanthocapsa sp. A, x200; (10) Diacanthocapsa brevithorax Dumitrica, x250; (11, 12) Diacanthocapsa antiqua (Squinabol), x200 (both); (13) Squinabollum fossile (Squinabol), x250; (14) Distylocapsa squama O’Dogherty, x200. Figs. (1-9,13)—Turonian; figs. (10-12,14) - Cenomanian.

Material: over 10 specimens.
Alievium sp. cf. A. superbum (Squinabol, 1914) Plate II, fig. 2
Remark. This form lacking spines is incompletely preserved. Nevertheless, the outer structure of shell and basal tubercles of lost spines are typical of the indicated species.
Distribution: lower Turonian of the western Cau¬casus.
Material: 2 specimens.
Multastrum sp. ex gr. M. regalis Vishnevskaya, 2001 Plate I, fig. 7
Hagiastridae gen. et sp. indet: Taketani, 1982, p. 50, pl. 10, fig. 3.
Multastrum sp.: Kurilov, 2005, Plate 32, fig. 19.; Vishnevskaya et al., 2005, Plate 13. fig. 5.
Material: 3 specimens.
Crucella messinae Pessagno, 1971 Plate II, fig. 4
Crucella messinae: Pessagno, 1971, p. 56, pl. 6, figs. 1-3; 1976, p. 32, pl. 1, fig. 4 (= holotype refig¬ured); Pessagno, 1977, p. 27, pl. 1, figs. 3, 4, 13; O’Dogherty, 1994, p. 368, pl. 70, figs. 21-24; Bragina, 2004, pl. 23, figs. 1, 2; pl. 38, fig. 5.
Non Crucella messinae: Taketani, 1982, p. 50, pl. 9, fig. 17 (= C. irwini Pessagno, 1971)
Distribution: lower Cenomanian of California; Albian-Cenomanian of Italy, Spain, and Atlantic Ocean; middle-upper Cenomanian of northern Turkey; upper Cenomanian-lower Turonian of the Crimean Mountains; uppermost Cenomanian-lower Turonian of the western Caucasus; undivided Coniacian-Campa- nian of the Russian plate.
Material: over 5 specimens.
Archaeocenosphaera ? mellifera O’Dogherty, 1994 Plate I, fig. 3
Hemicryptocapsa sp. A: Marcucci and Gardin, 1992, text-fig. 3. 1.
Archaeocenosphaera ? mellifera: O’Dogherty, 1994, p. 375, pl. 74, figs. 1-5; Salvini and Marcucci Passerini, 1998, Fig. 7.o; Bragina, 2004, pl. 14, fig. 3; pl. 36, fig. 14.
Distribution: Cenomanian-Campanian worldwide; middle Albian-lower Turonian of Italy and Spain; mid¬dle-upper Cenomanian of northern Turkey; upper Cen- omanian-lower Turonian of the Crimean Mountains; upper Cenomanian-lower Turonian of the western Caucasus; upper Cenomanian-lower Turonian, undi¬vided Coniacian-Campanian of the Russian plate.
Material: less than 10 specimens.
Phaseliforma sp. ex gr. P inflata Bragina, 2004
Plate I, fig. 6
Remarks. In shape and shell size, this form is similar to Phaseliforma inflata but has in distinction the quad¬rangle to heptagon knobs or relief projections.
Material: over 6 specimens.
Diacanthocapsa antiqua (Squinabol, 1903), emend. Bragina, 2004
Plate II, fig. 11,12
Theocorys antiqua: Squinabol, 1903, p. 135, pl. 8, fig. 25.
Diacanthocapsa antiqua: O’Dogherty, 1994, p. 220, pl. 36, figs. 25-28; Bragina, 2004, pl. 10, figs. 3, 7; pl. 31, figs. 11, 12; pl. 32, fig. 2.
Distribution: upper Albian-lower Turonian of Italy and Spain; upper Cenomanian of northern Turkey; upper Cenomanian-lower Turonian of the western Caucasus, lower Turonian of the Crimean Mountains.
Material: over 6 specimens.
Squinabollum fossile (Squinabol, 1903), emend. Bragina, 2004
Plate II, fig. 13
Clistosphaera fossilis: Squinabol, 1903, p. 130, pl. 10, fig. 11.
Squinabollum fossilis: Dumitrica, 1970, p. 83, pl. 19, figs. 118a-118c, 119a-119c; Teraoka and Kurimoto, 1986, pl. 4, fig. 4; Salvini and Marcucci Pas¬serini, 1998, Fig. 6.s; Kazintsova, 1993, p. 66, pl. XV, fig. 7.
Non Squinabollum fossilis: Vishnevskaya, 2001, pl. 22, figs. 7-11 (Sethocapsa?).
Squinabollum fossile: O’Dogherty, 1994, p. 203, pl. 32, figs. 4-10; Bragina, 2004, pl. 12, figs. 1-4.
Remark. Three or more sizeable spines in distal part of abdomen are oriented backward from cephalis.
Distribution: Albian-Santonian worldwide; upper Cenomanian of northern Turkey; lower Turonian of the Crimean Mountains; uppermost Cenomanian-lower Turonian of the western Caucasus; upper Santonian of the southwestern Sakhalin (Bragina, 2001).
Material: 6 specimens
Pseudodictyomitra pseudomacrocephala (Squinabol, 1903)
Plate II, fig. 7
Dictyomitra pseudomacrocephala: Squinabol,
1903, p. 139, pl. 10, fig. 2; Petrushevskaya and Kozlova, 1972, p. 550, pl. 2, fig. 5.
Pseudodictyomitra pseudomacrocephala: Pessa¬gno, 1977, p. 51, pl. 8, figs. 10, 11 (= specimens of Pes¬sagno, 1976, pl. 3, figs. 2, 3); Teraoka, Kurimoto, 1986, pl. 4, figs. 10, 11; O’Dogherty, 1994, p. 108, pl. 8, figs. 5-8; Salvini and Marcucci Passerini, 1998,
Fig. 8.h; Vishnevskaya, 2001, pl. 129, figs. 5, 9; Brag¬ina, 2004, pl. 7, fig. 4; pl. 32, figs. 9, 14-16.
Distribution: upper Albian-lower Turonian world-wide; middle-upper Cenomanian of northern Turkey; upper Cenomanian-lower Turonian of the Crimean Mountains; uppermost Cenomanian-lower Turonian of the western Caucasus.
Material: over 10 specimens.
Distylocapsa squama O’Dogherty, 1994, emend. Bragina, 2004
Plate II, fig. 14
Distylocapsa squama: O’Dogherty, 1994, p. 188, pl. 28, figs. 16-21; Salvini and Marcucci Passerini, 1998, Fig. 9.s; Bragina, 2004, pl. 4, fig. 6.
Distribution: upper Albian-lower Turonian of Italy and Spain; upper Cenomanian of northern Turkey; uppermost Cenomanian-lower Turonian of the western Caucasus; lower Turonian of the Crimean Mountains.
ACKNOWLEDGMENTS
The work was supported by the Russian Foundation for Basic Research, project no. 03-05-64964.
Reviewers A.S. Alekseev and V.A. Zakharov
REFERENCES
1. S. L. Afanas’ev, Geology of the Western Caucasus (Voentekhizdat, Moscow, 2004) [in Russian].
2. S. L. Afanas’ev and N. I. Maslakova, “Upper Cretaceous Deposits in the Northwestern Caucasus,” Tr. Vsesoyuzn. Zaochn. Politekh. Inst., Ser. Gidrogeol. Eng. Geol. 37, 106-136 (1967).
3. Yu. V. Agarkov, N. I. Boiko, and V. I. Sedletskii, Cherty Rocks of the Northern Caucasus and Potential Practical Use (Rostov. Gos. Univ., Rostov-on-Don, 1992) [in Rus¬sian].
4. A. H. Bouma, Sedimentology of Some Flysch Deposits. A Graphic Approach to Facies Interpretation (Elsevier, Amsterdam, 1962).
5. L. G. Bragina, “Cenomanian and Turonian Radiolarians from the Crimean Mountains,” Byull. Mosk. O-va Ispyt. Prir., Otd. Geol. 74 (3), 43-50 (1999).
6. L. G. Bragina, Extended Abstract of Candidate’s Disser¬tation in Geology and Mineralogy (GIN RAN, Moscow, 2001).
7. L. G. Bragina, “Cenomanian-Turonian Radiolarians of Northern Turkey and the Crimean Mountains,” Paleon- tol. J. 38 (Suppl. 4), 325-456 (2004).
8. L. G. Bragina and N. Yu. Bragin, “Stratigraphy and Radi- olarians from the Type Section of Perapedhi Formation (Upper Cretaceous of Cyprus),” Stratigr. Geol. Korre- lyatsiya 4 (3), 38-45 (1996) [Stratigr. Geol. Correlation 4 (3), 246-253 (1996)].
9. L. G. Bragina and N. Yu. Bragin, “Stratigraphy and Radi¬olarians of Upper Cretaceous Sedimentary Cover of the Arakapas Ophiolite Massif (Cyprus),” Stratigr. Geol.
Korrelyatsiya 14 (5), 506-6 (2006) [Stratigr. Geol. Cor-relation 14 (5), 507-523 (2006)].
10. P. Dumitrica, “Cryptocephalic and Cryptothoracic Nas- sellaria in some Mesozoic Deposits of Romania,” Rev. Roum. Geol. Geophys. Geoggr., Ser. Geol. 114 (1), 45¬124 (1970).
11. J. Erbacher, “Entwicklung und Palaoozeanographic mit- telkretazischer Radiolarien der westlichen Tethis (Ital- ien) und des Nordatlantiks,” Tubingen Mikropal. Mitt., No. 12, 1-119 (1994).
12. I. E. Karstens, The Ananuri Horizon (Gortoptekhizdat, Moscow, 1932) [in Russian].
13. L. I. Kazintsova, Radiolarians, Atlas of Guide Faunal Groups from the Cretaceous of Sakhalin (Nedra, St. Petersburg, 1993), [in Russian].
14. B. M. Keller, “Upper Cretaceous Flysch in the Western Caucasus,”. Tr. Inst. Geol. Nauk AN SSSR, Ser. Geol., No. 12, 21-27 (1940).
15. W. Kuhnt, J. Thurow, J. Wiedmann, and J. P. Herbin, “Oceanic Anoxic Conditions Around the Cenoma-nian/Turonian Boundary and the Response of the Biota,” Mitt. Geol. Palaontol., Hamburg Univ. 60, 205-246 (1986).
16. D. V. Kurilov, “New Localities of Jurassic and Creta-ceous Radiolarians in the Western Kamchatka,” in West¬ern Kamchatka: Geologic Development in the Mesozoic (Nauchnyi Mir, Moscow, 2005), pp. 55-76 [in Russian].
17. M. Marcucci and S. Gargin, “The Fosso Cupo Formation (Northern Latium, Italy): Redefinition and New Age Data from Radiolarian and Calcareous Nannofossil Bio¬stratigraphy,” Cretaceous Res. 13, 549-563 (1992).
18. D. P. Naidin and A. S. Alekseev, “Section of Cenomanian Deposits in the Kacha-Bodrak Interfluve (Crimea),” Izv. Vyssh. Uchebn. Zaved. Geol. Razved., No. 4, 11-25 (1980).
19. D. P. Naidin, A. S. Alekseev, and L. F. Kopaevich, “Fauna From Turonian Deposits in the Kacha-Bodrak Interfluve (Crimea) and the Cenomanian-Turonian Boundary,” in Organic Evolution and Biostratigraphy of the Mid-Cretaceous (Biologo-Pochv. Inst., Vladivostok, 1981), pp. 22-40 [in Russian].
20. D. P. Naidin and S. I. Kiyashko, “Geochemical Charac¬teristics of the Cenomanian-Turonian Boundary Depos¬its in the Crimean Mountains. Pap. 1. Lithology, Content of Organic Carbon and Some Elements,” Byull. Mosk. O-va Ispyt. Prir., Otd. Geol. 69 (1), 28-42 (1994a).
21. D. P. Naidin and S. I. Kiyashko, “Geochemical Charac¬teristics of the Cenomanian-Turonian Boundary Depos¬its in the Crimean Mountains. Pap. 2. Carbon and Oxy¬gen Isotope Composition; Accumulation Conditions of Organic Carbon,” Byull. Mosk. O-va Ispyt. Prir., Otd. Geol. 69 (2), 59-74 (1994b).
22. B. B. Nazarov and D. I. Vitukhin, “Methods for Separa¬tion of Fossil Radiolarians,” Izv. Akad. Nauk SSSR, Ser. Geol., No. 10, 95-101 (1981).
23. L. O’Dogherty, “Biochronology and Paleontology of Mid-Cretaceous Radiolarians from Northern Apennines (Italy) and Betic Cordillera (Spain),” Mem. Geol. Lau¬sanne, No. 21, 1-413 (1994).
24. E. A. Pessagno Jr., “Jurassic and Cretaceous Hagi- astridae from the Blake-Bahama Basin (Site 5 A, JOIDES Leg 1) and the Great Valley Sequence, Califor-
nia Coast Ranges,” Bull. Am. Paleontol 60 (264), 5-83 (1971).
25. E. A. Pessagno Jr., “Lower Cretaceous Radiolarian Bio-stratigraphy of the Great Valley Sequence and Fran-ciscan Coast Ranges,” Publ. Cusman Found. Foramin- iferal Res. 15, 1-87 (1977).
26. E. A. Pessagno Jr., “Radiolarian Zonation and Stratigra¬phy of Upper Cretaceous Portion of the Great Valley Sequence,” Micropaleontol. Spec. Publ. No. 2, 1-96 (1976).
27. M. G. Petrushevskaya and G. E. Kozlova, “Radiolaria,” in Initial Rep. Deep Sea Drill. Project, Vol. 14 (US Gov. Print. Off., Washington, 1972), pp. 495-648.
28. G. Salvini and M. Marcucci Passerini, “The Radiolarian Assemblages of the Bonarelli Horizon in the Umbria- Marche Apennines and Southern Alps, Italy,” Cretaceous Res. 19 (6), 777-804 (1998).
29. S. O. Schlanger, M. A. Arthur, H. C. Jenkins, et al., “The Cenomanian—Turonian Oceanic Anoxic Event. 1. Stratigraphy and Distribution of Organic Carbon-Rich Beds and the Marine 513C Excursion,” Spec. Publ. Geol., No. 26, 371-399 (1987).
30. S. Squinabol, “Le Radiolarie dei noduli selciosi nella Scaglia degli Euganei. Contribuzione I,” Riv. Ital. Pale- ontol. 9, 105-151 (1903).
31. S. Squinabol, “Radiolaria cretacee degli Euganei,” Atti Mem. Accad. Sci. Lett. Arti Padova, New. Ser. 20, 171-244 (1904).
32. S. Squinabol, “Contributo alla conoscenza dei Radiolarii fossili del Veneto. Appendice - Di Radiolari caratteris- tico del Secondario,” Mem. ist. geol. Univ. Padova 2, 249-306 (1914).
33. Y. Taketani, “Cretaceous Radiolarian Biostratigraphy of the Urakawa and Obira Areas, Hokkaido,” Sci. Rep. Tohoku Univ., 2nd Ser. (Geol.) 52 (1-2), 1-76 (1982).
34. Y. Teraoka and C. Kurimoto, “Cretaceous Stratigraphy of the Shimanto Terrane in the Uwajima Area, West Shikoku, Southwest Japan, with Reference to the Strati- graphic Distribution of Mega- and Radiolarian Fossils,” Bull. Geol. Surv. Japan 37 (8), 417-453 (1986).
35. N. B. Vassoevich, “Analogues of the Ananuri Formation (Lower Turonian) in the Southeastern Caucasus,” Dokl. Akad. Nauk SSSR 3 (4), 155-161 (1947).
36. V. S. Vishnevskaya, Jurassic-Cretaceous Radiolarian Biostratigraphy in Russia (GEOS, Moscow, 2001) [in Russian].
37. V. S. Vishnevskaya, Yu. V. Agarkov, G. S. Zakariadze, and K. M. Sedaeva, “Late Jurassic-Cretaceous Radiolar- ians from the Greater Caucasus as a Key for Deciphering Age and Formation Settings of Ophiolites in the Lesser Caucasus,” Dokl. Akad. Nauk 310 (6), 1417-1420 (1990).
38. V. S. Vishnevskaya, I. A. Basov, T. N. Palechek, and D. V. Kurilov, “Radiolarian and Foraminiferal Biostratigra¬phy of Jurassic-Cretaceous Deposits in the Western Kamchatka,” in Western Kamchatka: Geological Devel¬opments in the Mesozoic (Nauchnyi Mir, Moscow, 2005), pp. 6-54 [in Russian].
39. V. S. Vishnevskaya, T. N. Gorbachik, L. F. Kopaevich, and L. G. Bragina, “Foraminifers and Radiolarians across the Albian-Cenomanian and Cenomanian-Turo- nian Boundaries (Northern Peri-Tethys),” Stratigr. Geol. Korrelyatsiya 14 (5), 28-49 (2006) [Stratigr. Geol. Correlation 14 (5), 486-506 (2006)].
40. O. S. Vyalov, “Geological Investigations of 1931 in the Western Caucasus,” Zap. Vseros. Mineral. O-va, Ser. 2 63 (2), 27